Agro Sur Vol.25 (1) 62-80 1997
DOI: 10.4206/agrosur.1997.v25n1-08
CARACTERIZACIÓN DE SITIOS DE CRECIMIENTO DE SEIS
ESPECIES PRATENSES NATIVAS Y NATURALIZADAS DEL
DOMINIO HUMEDO DE CHILE1
Ignacio López C., Osear Balocchi L.,
Pablo Lailhacar M., Claudio Oyarzún S.
Universidad Austral de Chile
Facultad de Ciencias Agrarias
Instituto de Producción Animal
Casilla 567, Valdivia, Chile
1 Resultados de los Proyectos FONDECYT
92-0065 y 1951099
Recepción de originales: Mayo 16, de 1997
ABSTRACT
Characterization of the growing sites of six native
and naturalized pasture species in the Dominio Húmedo of Chile.
Key words: native forage species, naturalized
forage species, soil condition.
A large percentage of the grasslands of the Dominio Húmedo
of Chile is comprised of native and naturalized species. The presence and abundance
of a species in the field is the result of competition and the process of adaptation
to the environment. One of the ecosystemic elements which exerts selection pressure
on all of the species in the grassland is the soil, Therefore, the effect of
the soil on the species can modify the prairie, and the species that adapt best
can dominate. The objectives of the study were to characterize the sites where
the species that were studied dominated to determine the edaphic factors that
produced this selection and to verify if the environmental pressure of the soil
over the botanical composition resulted in selection of some of the species.
The species studied were Bromus valdivianus, Agrostis
capillaris, Lotus uliginosus, Holcus lanatus,
Anthoxanthum odoratum and Paspalum dasypleurus.
The zone studied was between Panguipulli and Puerto Montt. The species studied
dominated in the measured sites. The variables measured were slope (%), depth
of the soil (m), hydromorphism, texture, water pH (1:2.5), organic matter (%),
mineral nitrogen (ppm N-NO3), available phosphorus (ppm-Olsen), exchangeable
potassium (ppm), exchangeable sodium (meq/ 100 g.s.), exchangeable calcium (meq/100
g.s.), exchangeable magnesium (meq/100 g.s.), exchangeable bases (meq/100 g.s.),
exchangeable aluminum (meq/100 g.s.), aluminum saturation (%) and species presence
(%). A cluster statistical analysis was used to analize the data. Bromus
valdivianus dominated sites with well drained, medium textured, deep
soils and adequate levels of fertility. Agrostis capillaris
was the main species in flat to undulating sites with soils of medium depth,
heavy texture, slow drainage, high levels of exchangeable aluminum and high
aluminum saturation. Lotus uliginosus dominated depressed
sites with narrow soils, light texture, medium to deep permanent hydromorphism
and high levels of aluminum saturation and exchangeable aluminum. Holcus
lanatus was the principal species in flat to slightly depressed sites
with medium texture, medium depth and medium contents of aluminum. Anthoxanthum
odoratum dominated flat to undulated sites with high levels of aluminum
saturation and exchangeable aluminum. Paspalum dasypleurus
was dominant in depressed sites with medium stationary hydromorphism, medium
texture, medium depth of the soil, medium levels of aluminum saturation and
medium levels of exchangeable aluminum.
RESUMEN
Un alto porcentaje de las praderas del dominio Húmedo
de Chile están compuestas por especies naturalizadas. La presencia y
abundancia de las especies pratenses en una pradera son en gran parte el resultado
de un proceso de adaptación al medio y de competencia. Entre los elementos
ecosistémicos que ejercerían una presión de selección
sobre los componentes pratenses está el edafotopo. A partir de esto se
plantea que la acción del edafotopo sobre las especies de la pradera
podría llevar a que cada especie tendería a dominar cuando las
características y condición del sitio le fueran favorables.
Los objetivos planteados fueron caracterizar los sitios donde las especies en
estudio son dominantes, determinar los factores edáficos que inciden
en dicho desarrollo y verificar si la presión ambiental ejercida por
el edafotopo sobre las especies pratenses determinaban un crecimiento selectivo
de ellas.
Las especies estudiadas fueron Bromus valdivianus, Agrostis
capillaris, Lotus uliginosus, Holcus lanatus,
Anthoxanthum odoratum y Paspalum dasypleurus,
siendo la zona de muestreo tuvo como límite norte la ciudad de Panguipulli
y límite sur la de Puerto Montt. En los sitios muestreados dominaban
las especies deseadas. Las variables medidas fueron: pendiente (%), profundidad
de suelo (m), hidromorfismo, textura, pH al agua (1:2,5), materia orgánica
(%), nitrógeno mineral (ppm N-NO3), fósforo aprovechable
(ppm-Olsen), potasio intercambiable (ppm), sodio intercambiable (meq/100 g.s.),
calcio intercambiable (meq/100 g.s.), magnesio intercambiable (meq/100 g.s.),
suma de bases intercambiable (meq/100 g.s.), aluminio intercambiable (meq/100
g.s.), saturación de aluminio (%) y presencia de la especie (%). Para
analizar la información obtenida se utilizó el análisis
estadístico Cluster.
Bromus valdivianus dominó sitios profundos, de textura
media, con buen drenaje y con adecuados niveles de fertilidad. Agrostis
capillaris dominó sitios planos a ondulados, de profundidad
media, textura pesada, drenaje lento, altos niveles de aluminio intercambiable
y saturación de aluminio. Lotus uliginosus dominó
sitios depresionales, delgados, de textura liviana, con hidromorfismo permanente
medio a permanente profundo y altos niveles de saturación de aluminio
y de aluminio intercambiable. Holcus lanatus dominó
sitios planos a levemente depresionales, de texturas y profundidades medias
y con contenidos medios de aluminio. Anthoxanthum odoratum
dominó sitios planos a ondulados con altos niveles de saturación
de aluminio y de aluminio intercambiable. Paspalum dasypleurus
dominó en sitios depresionales con hidromorfismo estacional medio, textura
media, profundidad efectiva de suelo media y niveles medios de saturación
de aluminio y de aluminio intercambiable.
Todas las especies estudiadas dominaron en diferentes tipos de sitios. De manera
que la presencia de una u otra especie y su nivel de expresión en la
pradera está fuertemente relacionada con el tipo de sitio y la condición
del mismo. Finalmente las características edáficas más
determinantes para la dominancia de las especies fueron: la profundidad efectiva
de suelo, el tipo de hidromorfismo, la textura, la pendiente y los niveles de
aluminio intercambiable y de saturación de aluminio.
INTRODUCCIÓN
El Dominio Húmedo de Chile cuenta con 18.665.900 ha.
Se compone por una franja costera ubicada entre la Cordillera de la Costa y
el Océano Pacífico que se prolonga desde los 36° 15' LS hasta
la parte sur de la isla de Chiloé, denominándose provincia de
pastizales Húmeda de Verano Fresco o Valdiviana. Los territorios comprendidos
entre la Cordillera de la Costa y la Cordillera de los Andes, desde los 38°30'
LS hasta las zonas más australes del país pertenecen a la provincia
de pastizales Húmeda de Verano Fresco y Mésico o Los Lagos y la
provincia Húmeda de Verano Frío o Archipiélago Austral.
Además pertenece a este dominio la provincia Húmeda de Verano
Cálido o Isla de Pascua (Gastó et al.,
1987).
En este dominio predomina el clima templado lluvioso, especialmente
en su área meridional. Sin embargo, la extensión de Chile en latitud,
hace que la pluviometría y la temperatura varíen mucho de norte
a sur, situación que condiciona caracteres mediterráneos y submediterráneos
en ciertos sectores (Quintanilla, 1974).
Las praderas que se desarrollan en este dominio corresponden
en su mayor parte al tipo naturalizadas, de allí que poseen mucha importancia
para la ganadería de la zona. Los antecedentes que se disponen sobre
ellas, han evidenciado que con estas praderas se pueden obtener altos niveles
productivos, sin embargo la investigación que se ha hecho en ellas ha
sido limitada, de manera que aspectos básicos como son: adaptación
ambiental, fenología, productividad, entre otras se han mantenido dentro
de una caja negra.
La presente publicación pertenece a una importante
línea de trabajo que ha venido desarrollando el Instituto de Producción
Animal de la Universidad Austral de Chile tendiente a estudiar las especies
pratenses naturalizadas del dominio Húmedo de Chile. La hipótesis
planteada es que cada especie sería predominante en tipos de sitios particulares,
de manera que las características del sitio estarían ejerciendo
una acción ambiental sobre las especies de la pradera de manera que permitiría
un crecimiento selectivo de ellas.
Los objetivos fueron caracterizar los sitios donde las especies
en estudio dominan, determinar los factores edáficos que inciden en dicho
desarrollo y verificar si la presión ambiental ejercida por el edafotopo
sobre las especies pratenses determinaban un crecimiento selectivo de ellas.
MATERIALES Y MÉTODOS
La zona de muestreo comprendió praderas naturalizadas
de las provincias de pastizales Húmeda de Verano Fresco o Valdiviana
y Húmeda de Verano Fresco y Mésico o Los Lagos (Gastó
et al, 1987), teniendo como límite norte la ciudad
de Panguipulli y límite sur la de Puerto Montt.
Se estudiaron seis especies consideradas de gran importancia
como componentes de la pradera naturalizada del Sur de Chile. Estas fueron:
Bromus valdivianus, Agrostis capilletas,
Lotus uliginosus, Holcus lanatus, Anthoxanthum
odoratum y Paspalum dasypleurus.
Para cumplir con los objetivos, se seleccionaron dentro de
la zona mencionada diez sitios que debían estar dominados por cada una
de las especies en estudio. Una vez ubicados los sesenta sitios (diez para cada
especie), durante el período de primavera-verano, se muestreo y analizó
la pradera y el edafotopo. Las variables medidas fueron: pendiente (%), profundidad
de suelo (m), hidromorfísmo, textura, pH al agua (1:2,5), materia orgánica
(%), nitrógeno mineral (ppm N-NO3), fósforo aprovechable (ppm-Olsen),
potasio intercambiable (ppm), sodio intercambiable (meq/ 100 g.s.), calcio intercambiable
(meq/100 g.s.), magnesio intercambiable (meq/100 g.s.), suma de bases intercambiable
(meq/100 g.s.), aluminio intercambiable (meq/100 g.s.), saturación de
aluminio (%) y presencia de la especie (%).
Dentro de cada sitio se tomaron 5 submuestras de pradera con
el correspondiente suelo. El muestreo fue al azar. Para cada una de ellas se
obtuvo la disponibilidad instantánea de forraje, cortando a ras de suelo
la pradera contenida en un cuadrante de 0,04 m2, de manera que la superficie
total de pradera muestreada fue de 0,2 m2. Las muestras de suelo fueron tomadas
del suelo que estaba inmediatamente bajo la muestra de pradera y comprendieron
los primeros 20 cm de profundidad.
Las variables pendiente, exposición, textura, profundidad
de suelo e hidromorfísmo se clasificaron de acuerdo a Gastó
et al. (1993).
La composición botánica de la pradera fue obtenida
por medio de la separación manual de las diferentes especies en estado
fresco, en el Laboratorio de Forrajeras del Instituto de Producción Animal
de la Facultad de Ciencias Agrarias de la Universidad Austral de Chile, las
que luego fueron secadas en una estufa de aire forzado a 60°C por 48 hr.
Las muestras de suelo fueron analizadas en el Laboratorio de
Fertilidad de Suelos del Instituto de Suelos e Ingeniería Agrícola
de la Facultad de Ciencias Agrarias de la Universidad Austral de Chile.
Los datos de cada variable en los diez sitios de cada especie,
fueron analizados por medio del diseño estadístico Cluster (conglomerados),
utilizando el paquete estadístico SPSS.
RESULTADOS Y DISCUSIÓN
Análisis de la fitocenosis.
En el Cuadro 1 se observa la composición
botánica promedio de los diferentes sitios analizados, los que se encontraban
fuertemente dominados por las especies estudiadas.
| Cuadro 1. |
Composición botánica (% BMS) de los
sitios evaluados. |
| |
Botanical composition (% DM) of the evaluated sites. |
|
| Especies |
Sitios de crecimiento |
| (% BMS) |
|
| |
B. valdivianus |
A. capillaris |
L. uliginosus |
H. lanatus |
A. odoratum |
P. dasypleurus |
|
B. valdivianus |
79,4 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
|
A. capillaris |
0,7 |
83,0 |
3,4 |
7,6 |
11,8 |
5,4 |
|
L. uliginosus |
0,0 |
0,2 |
79,0 |
1,1 |
0,6 |
1,0 |
|
H. lanatus |
0,6 |
2,7 |
4,1 |
84,7 |
1,7 |
4,0 |
|
A. odoratum |
0,0 |
6,5 |
0,5 |
0,6 |
80,1 |
1,1 |
|
P. dasypleurus |
0,0 |
0,0 |
1,4 |
0,0 |
0,0 |
78,0 |
|
L. perenne |
12,1 |
0,0 |
0,1 |
4,9 |
0,0 |
0,4 |
|
T. repens |
0,1 |
0,0 |
1,7 |
0,7 |
0,0 |
0,2 |
|
Otras grams. |
6,2 |
0,0 |
0,3 |
0,0 |
2,5 |
0,3 |
|
Otras legs. |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
|
Juncaceae y
Rununculaceae
|
0,0
|
0,2
|
7,6
|
0,1
|
0,0
|
6,4
|
|
Esp. de
Hoja Ancha
|
0,9
|
7,4
|
1,9
|
0,3
|
3,3
|
3,2
|
|
Además de analizar los porcentajes de las especies en
estudio es importante observar cuales eran las especies acompañantes
y en los niveles en que ellas se encontraban presentes, ya que son indicativas
de una condición de pradera determinada y tienen un claro papel ecológico-productivo.
Estos conceptos son expresados por diversos autores como Dyksterhuis
(1959) y Gastó et al. (1993).
En los sitios de crecimiento de B. valdivianus
este presentó 79% (Cuadro 1) de presencia en peso seco.
La principal especie competidora por los nichos ecológicos fue L
perenne (12 %). D. glomerata se constituyó
como especie compe-tidora terciaria (6%). Estas tres especies se las encuentra
frecuentemente compitiendo en las praderas de la zona, estableciéndose
en los suelos de mayor potencial productivo (Cuevas, 1980;
Cuevas et al, 1982; Bernier,
1985).
A. capillaris presentó una dominancia
de 83%, teniendo como especie competidora a A. odoratum (2,7%).
En estos sitios las especies de hoja ancha (7,4%) y A.odoratum
tienen un papel de indicadoras del grado degradación del sitio. Si bien
tienen un limitado papel productivo, estas especies corresponden a los últimos
eslabones de la degradación de las praderas, y son importantes porque
crecen bajo condiciones extremas donde otras especies de mayor producción
y requerimientos no son capaces de establecerse.
L. uliginosus alcanzó 79%, encontrándose
más frecuentemente: H. lanatus (4,1 %), A.
capillaris (3,4%) y Juncaceas y Ciperaceas (7,6%). Estas tres últimas
se asocian continuamente a L. uliginosus, indicando una situación
de hidromórfica, lo que ocurre frecuentemente en suelos ñadis
(Montaldo y Pessot, 1974; Ramirez et
al., 1990; Ramírez et al,
1991).
H. lanatus tuvo 84.7% de dominancia, subdominantes
fueron L. perenne (4,9%) y A. capillaris
(7,6%). Esto está indicando que H. lanatus tiene la
capacidad de desarrollarse en una amplia gama de sitios de características
medias de fertilidad ocupando sitios intermedios entre los de baja fertilidad,
dominados normalmente por A. capillaris y los de alta fertilidad
dominados por L. perenne. Esta capacidad colonizadora de H.
lanatus también es comentada por Grime et
al. (1989)
Los sitios dominados por A. odoratum tuvieron
80% de esta especie, siendo A. capillaris (11,8%) la especie
que le competía más fuertemente y capaz de adaptarse a las condiciones
de sitio en las que predomina A. odoratum. Otra especie que
acompañó a A. odoratum fue Cynosorus
echinatus pero en menor intensidad. Grime et
al. (1989) indican que esta especie se asocia a a ecosistemas pratenses
con niveles de producción bajos a medios y sujetos a leves a medios niveles
de perturbación. Grime (1979) define perturbación
como el mecanismo por el cual se limita el crecimiento de la planta y que causa
una parcial o total destrucción de la biomasa. Este fenómeno está
asociado a la actividad de herbívoros, patógenos y artrópica
y a la acción del viento, bajas temperaturas, erosión, incendios
y sequía.
P. dasypleurus (78%)) tuvo como especies
acompañantes a A. capillaris (5 A%\ J.procerus
(6,4%) y H. lanatus (4,0%), situación similar, desde
el punto de vista de las especies acompañantes, a la encontrada con L.
uliginosus. Esto podría sugerir que P. dasypleurus
y L. uliginosus se desarrollarían en edafotopos similares,
sin embargo, su presencia en los sitios como especie acompañante fue
para ambos casos cercano al 1%, lo que indicaría que existen factores
que impiden el desarrollo de ellas bajo dichas situaciones. Ello se analizará
más adelante.
Análisis de las variables edáficas evaluadas.
Del análisis estadístico de todas las variables
en conjunto, se observó que en general la dominancia de las especies
estaba explicada en 83% por un lado debido a la interacción entre aluminio
intercambiable y la saturación de aluminio, y por otro lado a la interacción
de las variables profundidad, pendiente, textura e hidromorfísmo.
Lo anterior indicaría que entre las variables de fertilidad
medidas, los contenidos de aluminio del suelo determinarían el nivel
de dominancia de una u otra especie, en conjunto con las variables físicas
que definen los diferentes tipos de sitios (profundidad, textura e hidromorfísmo)
más la pendiente, determinada por la geoforma. Esto coincide con lo indicado
por Gastó et al.(l993) al definir los conceptos
de distrito y sitio, siendo el primero definido por la pendiente (escala de
resolución 1:250.000) y el segundo por la interacción entre textura,
profundidad e hidromorfísmo (escala de resolución 1:10.000), todos
constituyentes permanentes del suelo.
Al analizar sólo los factores físicos del edafotopo,
las variables que tuvieron una alta correlación fueron por una parte
la profundidad con el hidromorfísmo y por otra la pendiente con la textura,
explicando ellas en un 76% la dominancia de las especies.
Las variables de fertilidad más relevantes fueron agrupadas
de la siguiente manera, por un lado pH, aluminio intercambiable y saturación
de aluminio y por otro el potasio intercambiable, calcio intercambiable, magnesio
intercambiable y la suma de bases intercambiable, explicando de esta forma en
un 77% la dominancia de la especies.
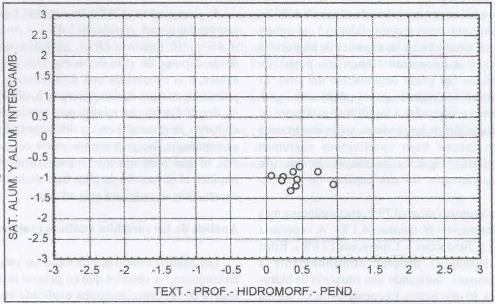 |
| |
| Figura 1. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por B.
valdivianus.
Distribution of the most relevant edaphic variables for B. valdivianus
dominated sites. |
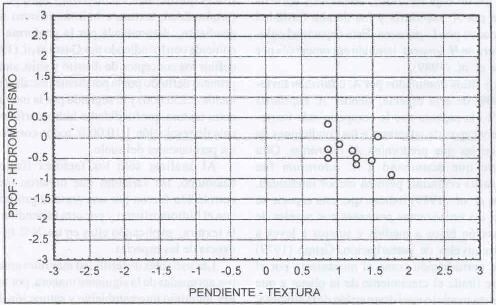 |
| |
| Figura 2. |
Distribución de las variables
físicas del suelo más relevantes para los sitios dominados
por B. valdivianus.
Distribution of the most relevant soil physical variables for B. valdivianus
dominated sites. |
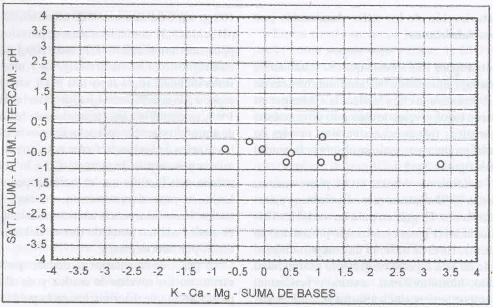 |
| |
| Figura 3. |
Distribución de las variables
de fertilidad de suelo más relevantes para los sitios dominados
por B. valdivianus.
Distribution of the most relevant soil fertility variables for B.
valdivianus dominated sites. |
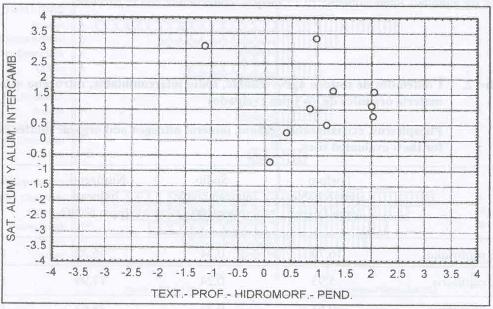 |
| |
| Figura 4. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por A.
capillaris.
Distribution of the most relevant edaphic variables for A. capillaris
dominated sites. |
Caracterización de los sitios dominados por
Bromus valdivianus.
En la Figura 1 se aprecia que B.
valdivianus domina bajo situaciones bastante homogéneas de suelo.
Presenta una clara tendencia a dominar en suelos con bajos niveles de aluminio
y que por sus características físicas poseerían altos niveles
de potencial productivo, como se observa claramente en las figuras 2
y 3.
B. valdivianus domina sitios planosuaves,
profundos (0,93m), textura media (franco) y drenaje moderado. El gráfico
para las variables físicas estudiadas (Figura 2) ratifica
lo presentado en la Figura 1. En él se observa un comportamiento
definido de esta especie respecto de profundidad de suelo, hidromorfísmo,
textura y pendiente. Este comportamiento indicaría que B. valdivianus
sería poco tolerante a las variaciones de las variables físicas
que caracterizan a los sitios, es decir, poseería una baja plasticidad,
lo que ecológicamente constituye una desventaja, en especial a lo que
se refiere a una menor capacidad de colonización de nuevos sitios.
Desde el punto de vista de la fertilidad de los suelos, B.
valdivianus domina sitios con bajos niveles de aluminio intercambiable
(0,15 meq/ 100 g. s.), baja saturación de aluminio (2,72%), pH ácido
(5,5), contenidos altos de potasio (243,5 ppm), medios de calcio (5,76 meq/100
g. s.), altos en magnesio (1,81 meq/100 g. s.) y medios en suma de bases (6,54
meq/100 g. s.), siendo esta especie, de acuerdo con los cluster de las Figuras
1 y 3, susceptible a las variaciones de
los niveles de aluminio y acidez del suelo. Esto no ocurre con los niveles de
las bases, frente a los cuales tolera un margen amplio de variación,
por lo que sería mucho más factible que el estrés, y por
ende la limitante para el crecimiento, en este tipo de suelos, lo constituyera
la cantidad de aluminio en el medio edáfico antes de que se debiera a
una estrés por falta de bases.
A partir de lo anterior se deduce que un aumento en los niveles
de acidez y de aluminio provocarían una disminución en la población
de B. valdivianus ingresando especies tolerantes como A.
capillaris y A. odoratum.
Por otra parte, B. valdivianus en suelos
que poseen mejores niveles de fertilidad, y por lo tanto menores tenores de
acidez y de aluminio, tiende a decrecer su presencia en la pradera, debido al
ingreso de especies más agresivas y altamente competitivas por los recursos
lumínicos y edáficos, como ocurre con L perenne.
| Cuadro 2. |
Contenidos de fósforo
aprovechable, sodio intercambiable, nitrógeno mineral y materia
orgánica de los sitios evaluados. |
| |
Phosphorus, exchangeable sodium, mineral
nitrogen and organic matter content for thew evaluated sites. |
| |
|
Fósforo |
Sodio |
Nitrógeno |
Materia |
| |
|
aprovechable |
intercambiable |
mineral |
orgánica |
| |
|
(ppm-Olsen) |
(meq/100g) |
(ppm N-NO3) |
(%) |
| |
|
| |
B. valdivianus |
16,70 |
0,34 |
36,22 |
18,39 |
| |
|
| |
A. capillaris |
5,93 |
0,24 |
17,99 |
12,08 |
| |
|
| |
L. uliginosus |
9,63 |
0,27 |
38,63 |
19,51 |
| |
|
| |
H. lanatus |
10,14 |
0,41 |
38,64 |
17,83 |
| |
|
| |
A. odoratum |
5,39 |
0,36 |
19,04 |
16,74 |
| |
|
| |
P. dasypleurus |
5,11 |
0,28 |
30,31 |
15,86 |
| |
|
 |
| |
| Figura 5. |
Distribución de las variables
físicas del suelo más relevantes para los sitios dominados
por A. capillaris.
Distribution of the most relevant soil physical variables for A. capillaris
dominated sites. |
 |
| |
| Figura 6. |
Distribución de las variables
de fertilidad del suelo más relevantes para los sitios dominados
por A. capillaris.
Distribution of the most relevant soil fertility variables for A.
capillaris dominated sites. |
 |
| |
| Figura 7. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por L.
uliginosus.
Distribution of the most relevant edaphic variables for L. uliginosus
dominated sites. |
 |
| |
| Figura 8. |
Distribución de las variables
físicas del suelo más relevantes para los sitios dominados
por L. uliginosus.
Distribution of the most relevants soil physical variables for L.
uliginosus dominated sites. |
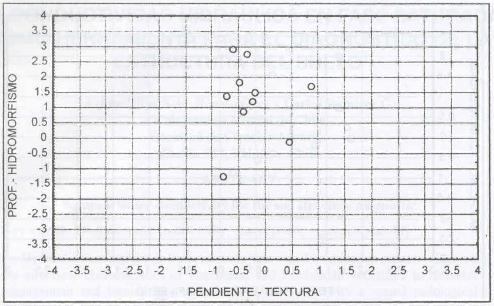
Caracterización de los sitios dominados por Agrostis
capillaris.
Esta es una especie que tendría una mayor capacidad
de adaptación a diversos ambientes que B. valdivianus,
debido a que posee una distribución más amplia en el espectro
de las variables estudiadas (Figura 4). Además esto
significa que es capaz de tolerar condiciones de suelo con mayor estrés,
como son el alto contenido de aluminio en el suelo, texturas arcillosas, fuertes
pendientes, etc.
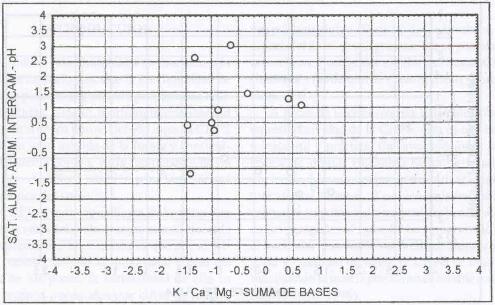
En la Figura 6 se observa que A. capillaris
prospera en suelos ácidos, con altos tenores de aluminio y pobres en
bases. Cuando la condición de fertilidad del suelo mejora, A.
capillaris tiende a bajar su presencia en la pradera, posiblemente
porque tiene que competir con especies más agresivas que se desarrollan
normalmente bajo mejores condiciones de fertilidad. Por lo tanto su abundancia
en la pradera se debería más a un fenómeno de tolerancia
que de competencia.
Esta especie domina en sitios plano-inclinados a suavemente
ondulados, de profundidad media (0,45 m), textura pesada (arcillo francesa a
arcillo arenosa) y drenaje lento (Figura 5).
Montaldo (1975) indica que A.
capillaris es una especie que se ha adaptado a numerosos ecosistemas
de la zona sur, no siendo para ella un factor limitante el hidromorfismo existente.
A. capillaris domina sitios con altos contenidos
de aluminio intercambiable (1,25 meq/100 g.s.), alta saturación de aluminio
(65,36%), pH fuertemente ácido (5,2), tenores altos de potasio (152,9
ppm), bajos en calcio (1,459 meq/100 g. s.), bajos en magnesio (0,54 meq/100
g. s.) y baja suma de bases (2,63 meq/100 g. s.). Por lo tanto son suelos que
se encuentran en una pobre condición de fertilidad y que, desde este
punto de vista, poseen fuertes restricciones para el desarrollo de especies
de alto potencial productivo como L. perenne, siendo este
justamente el valor ecológico y productivo de A. capillaris.
Ramirez et al. (1991 b) indican que A.
capillaris se adapta a suelos con pH entre 4,5 y 6,5, ubicándose
de preferencia en los suelos ácidos.
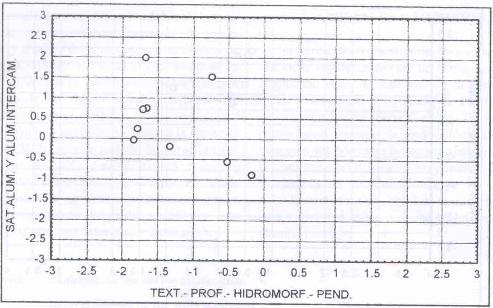
Caracterización de los sitios dominados por
Lotus uliginosus.
El diagrama del resultado del cluster efectua do para esta
especie indica que en general esta especie tiende a dominar en sitios depresionales,
de escasa profundidad, de textura liviana y con altos contenidos de aluminio
(Figura 7).
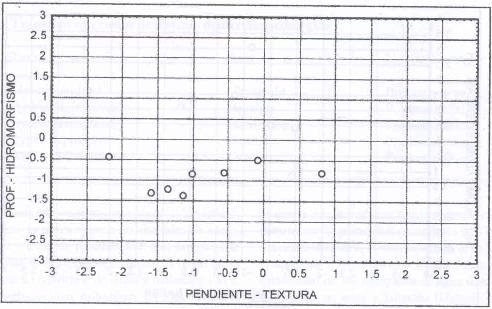
Esta especie domina en sitios depresionales, delgados (0,38
m), de textura liviana (franco arenosa), hidromorfismo permanente medio a permanente
profundo (Figura 8).
Ramirez et al. (1991)
describen el valor indicador ecológico para diferentes especies pratenses,
respecto de una serie de variables entre las cuales se encuentra la humedad
del medio edáfico. Los autores concluyen que B. valdivianus
se ubica entre las especies indiferentes a los niveles hídricos, al igual
que A. capillaris, mientras que a L. uliginosus
se la clasifica dentro de las plantas de suelos húmedos.
L. uliginosus domina sitios con altos contenidos
de aluminio intercambiable (0,71 meq/100 g. s.), alta saturación de aluminio
(19,69%), pH fuertemente ácido (5,2), contenidos altos en potasio (161,7
ppm), medios en calcio (3,35 meq/ 100 g. s.), medios en magnesio (0,91 meq/100
g. s.) y media-baja suma de bases (4,96 meq/100 g. s.).
De lo anterior se desprende que las condiciones de fertilidad
de suelo en que se desarrolla L. uliginosus son similares
a las que tienen los sitios dominados por A. capillaris ,
sin embargo, esta última posee mayores rangos de tolerancia a los contenidos
de aluminio (Figura 7 y Figura 9).
La diferencia entre los sitios dominados L. uliginosus
y A. capillaris radica principalmente en las características
físicas de ellos, especialmente a lo que se refiere a pendiente, textura
e hidromorfismo. Por lo tanto, L. uliginosus corresponde a
una especie altamente tolerante a los altos niveles de hidromorfismo, típicos
de los sitios depresionales con mal drenaje, sin embargo es altamente susceptible
a los déficit hídricos estivales, de manera que debido a esta
razón no competiría por los sitios de crecimiento de A.
capillaris.
Grime et al. (1989) indican
que L. uliginosus coloniza sitios depresionales pero que no
correspondan a sitios permanentemente sumergidos (hidromorfismo permanente superficial).
Además no se presenta en suelos con pH menores a 4,5, siendo común
en suelos con pH 4,5 a 6,5.
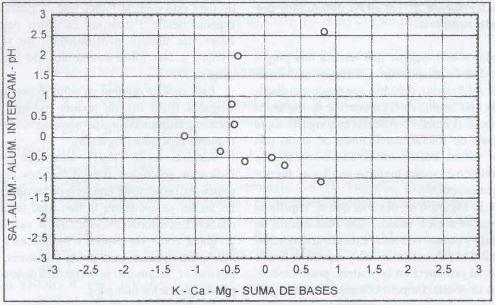 |
| |
| Figura 9. |
Distribución de las variables
de fertilidad de suelo más relevantes para los sitios dominados
por L. uliginosus.
Distribution of the most relevant soil fertility variables for
L. uliginosus dominated sites. |
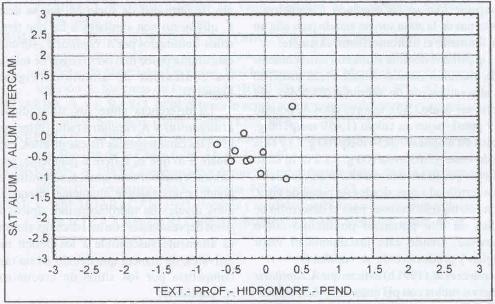 |
| |
| Figura 10. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por H.
lanatus.
Distribution of the most relevant edaphic variables for H. lanatus
dominated sites. |
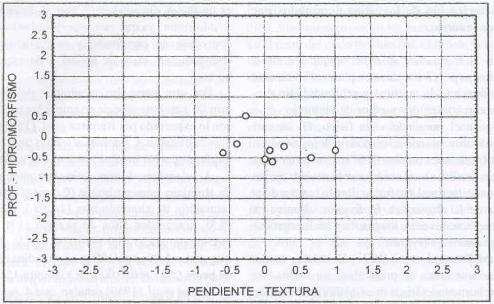 |
| |
| Figura 11. |
Distribución de las variables
físicas del suelo más relevantes para los sitios dominados
por H. lanatus.
Distribution of the most relevant soil physical variables for H. lanatus
dominated sites. |
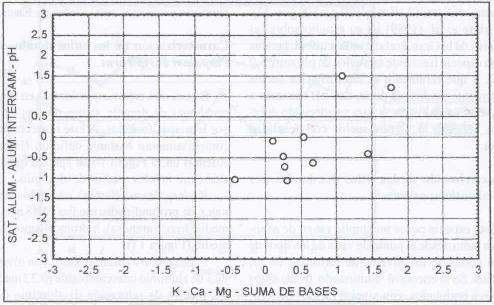 |
| |
| Figura 12. |
Distribución de las variables
de fertilidad de suelo más relevantes para los sitios dominados
por H. lanatus.
Distribution of the most relevant soil fertility variables for H. lanatus
dominated sites. |
Caracterización de los sitios dominados por
Holcus lanatus.
En la Figura 10 se observa que los sitios
dominados por H. lanatus son planos a levemente depresionales,
de texturas y profundidades medias y con contenidos medios de aluminio.
Desde el punto de vista físico, H. lanatus
domina sitios planos a levemente depresionales, de profundidades medias (0,67
m), textura media (franco arcillosa), con hidromorfísmo estacional medio
a estacional profundo. Por lo tanto a diferencia de L. uliginosus,
H. lanatus domina en sitios ecotonales entre los planos y
los fuertemente depresionales (Figura 11).
Para Ramirez et al. (1991)
H. lanatus es una especie que crece de preferencia en suelos
con mayor humedad. Grime et al. (1989)
indican que es una especie típica que coloniza suelos con problemas de
drenaje.
Esta especie domina en sitios de niveles medios de fertilidad,
caracterizados por tener niveles medios de aluminio intercambiable (0,4 meq/100
g.s.), saturación de aluminio medio (5,87%), pH ácido (5,4), altos
contenidos de potasio (257,9 ppm), niveles medios de calcio (4,82 meq/100 g.
s.), altos de magnesio (1,3 meq/100 g. s.) y medio-alta suma de bases (7,19
meq/100 g. s.) (Figura 12).
Grime et al. (1989) en
su estudio sobre las praderas de la Gran Bretaña indica que H.
lanatus es una especie frecuente en suelos de pH entre 5,0 y 6,0, y
que raramente se desarrolla en suelos fuertemente ácidos, es decir, con
pH menores a 4,8. Además esta especie para su adecuado desarrollo necesita
al menos suelos con fertilidad media.
Caracterización de los sitios dominados por
Anthoxanthum odoratum.
Esta especie posee un amplio rango de adaptación tanto
desde el punto de vista de los tipos de sitios como de los niveles de fertilidad
de los mismos. Se la encontró dominando desde sitios planos a ondulados,
con niveles medios de fertilidad (Figura 13).
Domina sitios planos a ondulados (6,8% de pendiente), de profundidades
medias a profundas (0,67 m), textura pesada (arcillo arenosa) y drenaje lento
a moderado (Figura 14).
Para Grime et al. (1989) la pendiente no
es un factor discriminante para el desarrollo de A. odoratum
, ya que es capaz de colonizar sitios con cualquier pendiente, y es particularmente
abundante en sitios de solana o de exposición norte.
Por otra parte, los resultados del presente estudio para esta
especie no coinciden totalmente con lo expresado por Ramirez
et al. (1991), quienes clasifican a A. odoratum
como una especie indiferente a los niveles hídricos del suelo.
A. odoratum domina sitios con altos niveles
de aluminio intercambiable (0,6 meq/100 g. s.), saturación de aluminio
alta (14,1%), pH ácido (5,5), contenidos altos de potasio (171 ppm),
medios de calcio (3,9 meq/100 g. s.), altos de magnesio (1,06 m eq/100 g. s.)
y medios en suma de bases (5,77 meq/100 g. S.) (Figura 15).
Grime et al. (1989) señalan
que A. odoratum se desarrolla frecuentemente en sitios de
pH 4,5 a 6,0.
Del análisis de las características de los sitios
dominados por A. odoratum se desprende que se desarrolla en
sitios muy similares a los de A. capillaris, especialmente
en lo que se refiere a las características físicas de los sitios,
sin embargo la diferencia radica en que A. odoratum domina
en sitios no tan ácidos, con menores tenores de aluminio y con mayores
niveles de bases.
Caracterización de los sitios dominados por
Paspalum dasypleurus.
Esta es otra especie que domina en suelos con problemas de
drenaje, como son los generados por las napas freáticas. A este respecto
posee un comportamiento bastante definido (Figura 16). Además
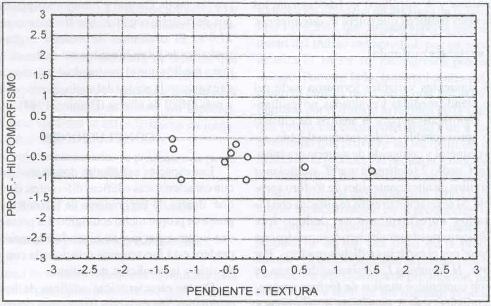
en la Figura 16 se aprecia que domina sitios con niveles
medios de aluminio.
P. dasypleurus domina en suelos depresionales,
de profundidades medias (0,48 m), textura media (franco arenosa), hidromorfísmo
estacional medio (Figura 17).
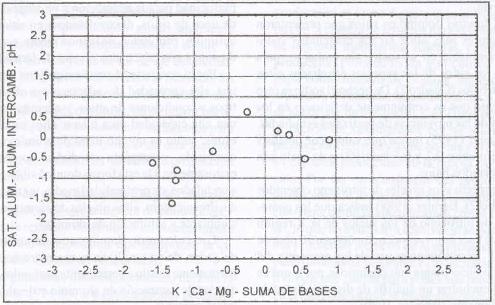
P. dasypleurus domina sitios con niveles
medios de aluminio intercambiable (0,32 meq/100 g. s.), medios de saturación
de aluminio (9,84%), pH ácido (5,45), altos contenidos de potasio (162
ppm), medio-bajo niveles de calcio (2,82 meq/ 100 g. s.), medios de magnesio
(0,81 meq/100 g. s.) y suma de bases medio-bajo de 4,32 meq/100 g. s. (Figura
18).
La diferencia entre los sitios dominados por A. odoratum
y A. capillaris es básicamente los altos niveles freáticos
que tolera P. dasypleums.
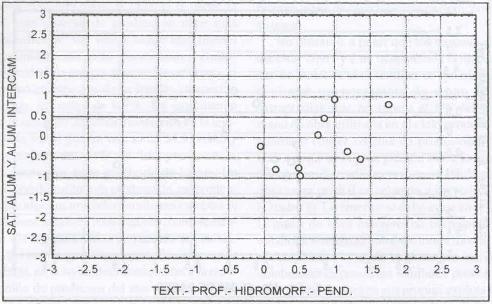 |
| |
| Figura 13. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por A.
odoratum.
Distribution of the most relevant edaphic variables for A. odoratum
dominated sites. |
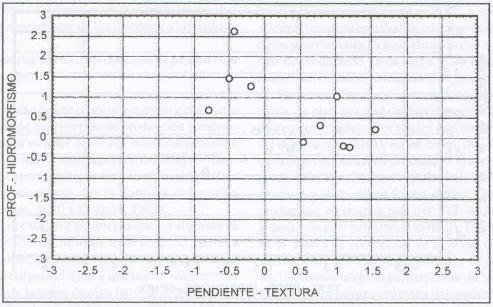 |
| |
| Figura 14. |
Distribución de las variables
edáficas más relevantes para los sitios dominados por A.
odoratum.
Distribution of the most relevant soil physical variables for A. odoratum
dominated sites. |
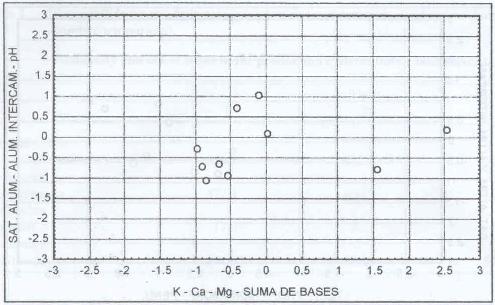 |
| |
| Figura 15. |
Distribución de las variables
de fertilidad de suelo más relevantes para los sitios dominados
por A. odoratum.
Distribution of the most relevant soil fertility variables for A.
odoratum dominated sites. |
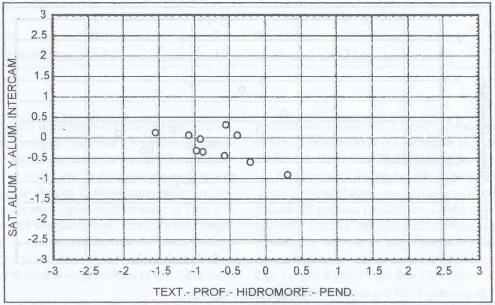 |
| |
| Figura 16. |
Distribución de las variables edáficas
más relevantes para los sitios denominados P. dasypleurus
Distribution of the most relevant edaphic variables
for P. dasypleurus dominates sites.
|
 |
| |
| Figura 17. |
Distribución de las variables físicas del suelo
más relevantes para los sitios denominados por P.
dasypleurus.
Distribution of the most relevant soil physical variables for
P. dasypleurus dominated sites.
|
 |
| |
| Figura 18. |
Distribución de las variables de fertilidad
del suelo más relevantes para los sitios dominados por
P. dasypleurus.
Distribution of the most relevant soil fertility variables for
P. dasypleurus dominated sites.
|
Otras variables evaluadas.
Las siguientes variables formaron parte del estudio, siendo
medidas y evaluadas, no resultando ser significativas para el análisis
de cluster: fósforo aprovechable, sodio intercambiable, nitrógeno
mineral y contenido de materia orgánica.
En el Cuadro 2 se observa que B.
valdivianus domina sitios de altos contenidos de fósforo aprovechable.
Si se relacionan estos resultados con los entregados anteriormente se deduce
que B. valdivianus puede ser un claro competidor con L. perenne
por el espacio en dichas praderas. Por otra parte, H. lanatus
y L. .uliginosus dominan en sitios con contenidos medios de
fósforo aprovechable, mientras que A. capillaris, A.
odoratum y P. dasypleums dominan en sitios con bajos
niveles de este elemento.
Pinochet (1990) señala que suelos
con contenidos menores a 8 ppm de P-Olsen, en suelos muestreados a 20 cm de
profundidad, serían deficitarios de este elemento. En su estudio indica
que el fósforo es un nutriente que es deficitario normalmente en los
suelos de la provincia de Valdivia.
H. lanatus dominó en sitios que presentaron
los valores más altos en los contenidos sodio intercambiable en el suelo,
sin embargo, para todos los sitios de las especies estudiadas estos fueron bajos
(Cuadro 2). De hecho el sodio es otro elemento que es comúnmente
deficitario en los suelos de las provincias de pastizales estudiadas. Rodríguez
(1993) indica que valores de análisis de suelo entre 0,2 y 0,5 meq/100
g de suelo son considerados bajos.
Respecto a los niveles de nitrógeno obtenidos (Cuadro
2), Bernier (1988) indica que los contenidos de nitrógeno
de los suelos de la X región tienden a ser bajos, es decir, menores a
20 ppm de N-NO3. El nitrógeno es uno de los nutrientes del
sistema suelo-planta más dinámicos, por lo que es difícil
encontrar un análisis de disponibilidad de este elemento (Bernier,
1980).
Para todas las especies los contenidos de materia orgánica
del suelo fueron altos (Cuadro 2), encontrándose dentro
de los rangos esperados para praderas naturalizadas de la zona. Al respecto
Sierra (1991) señala que los suelos de las provincias
de Osorno y Llanquihue poseen rangos de materia orgánica que fluctúan
entre 8,0 y 21,0%. El contenido de materia orgánica del suelo incide
sobre la acidez del suelo, de manera que a medida que el contenido de materia
orgánica es mayor, la acidez del suelo es más persistente y más
difícil de alterar (Bernier, 1984).
CONCLUSIONES
Las especies estudiadas dominaron en sitios con características
edáficas diferentes, de manera que dentro de las praderas de la zona
cada una posee su propio nicho ecológico. La presencia de una u otra
especie y su nivel de expresión en la pradera está fuertemente
relacionada con el tipo de sitio y la condición del mismo.
De las características edáficas de los sitios
estudiados, las variables físicas más determinantes para la dominancia
de las especies fueron la profundidad efectiva de suelo, el tipo de hidromorfísmo,
la textura y la pendiente, y entre las químicas fueron los tenores de
aluminio intercambiable y el nivel de la saturación de aluminio.
Bromus valdivianus es una especie que dominó
en sitios de alta fertilidad y alto potencial productivo y no sería una
especie con una alta plasticidad para la adaptación a un amplio rango
de tipos de sitios, desarrollándose en sitios bien definidos: profundos,
de textura media, con buen drenaje y con adecuados niveles de fertilidad.
Por el contrario, Agrostis capillaris presentó
una alta capacidad de adaptación a diferentes tipos y condiciones de
sitios, y además presentó una alta capacidad para tolerar altos
niveles de estrés, como es un alto nivel de aluminio intercambiable,
y competir en dichas condiciones exitosamente. De esta forma dominó sitios
planos a ondulados, de profundidad media, textura pesada, drenaje lento, altos
niveles de aluminio intercambiable y saturación de aluminio.
Lotus uliginosus dominó sitios depresionales,
delgados, de textura liviana, con hidromorfismo permanente medio a permanente
profundo y altos niveles de saturación de aluminio y de aluminio intercambiable.
Holcus lanatus se presentó como una
especie capaz de desarrollarse en sitios de alto potencial productivo de praderas,
pero donde la condición edáfica estaba levemente desmedrada. Dominó
sitios planos a levemente depresionales, de texturas y profundidades medias
y con contenidos medios de aluminio.
Anthoxanthum odoratum fue otra especie que
presentó un amplio rango de adaptación edáfica dominando
sitios planos a ondulados con altos niveles de saturación de aluminio
intercambiable.
Paspalum dasypleunis dominó en sitios
depresionales con hidromorfismo estacional medio, textura media, profundidad
efectiva de suelo media y ni veles medios de saturación de aluminio y
de aluminio intercambiable.
BIBLIOGRAFÍA
BERNIER, R. 1980. Avances en fertilidad de
suelos de la Xa Región. INIA, Estación Experimental Remehue. Boletín
Técnico (Chile) N° 40. 27 p.
BERNIER, R. 1984. Efectos de la aplicación
de cal en el suelo. INIA, Estación Experimental Remehue. Boletín
Técnico (Chile) N° 79. 15 p.
BERNIER, R. 1985. Fertilización de
praderas. III. Tipos de praderas de la Xa Región. INIA, Estación
Experimental Remehue. Boletín Técnico (Chile) N° 90. 13 p.
BERNIER, R. 1988. Fertilización de
praderas. INIA, Estación Experimental Remehue. Boletín Técnico
(Chile) N° 136. 18 p.
CUEVAS, E. 1980. Producción, manejo
y utilización de praderas. In..: L. Latrille, ed. Segundo Curso Nacional
de Producción de Leche. Universidad Austral de Chile, Facultad de Ciencias
Agrarias, Instituto de Producción Animal. Serie B-4 Tomo I. Valdivia,
Chile, pp. 75-114.
CUEVAS, E., O. BALOCCHI, R. ANRIQUE y M. JORQUERA.
1982. Valor nutritivo de las principales especies de una pradera permanente
de la Décima Región. I. Proteína y pared celular. Agro
Sur (Chile) 10: 2 - 6.
DYKSTERHUIS, E. 1959. Range conservation based
on sites and condition classes. Journal of Soil and Water Conservation. 13:
104 - 115.
GASTO, J., S. GALLARDO y D. CONTRERAS. 1987.
Caracterización de los pastizales de Chile. Reinos, Dominios y Provincias.
Sistemas en Agricultura. Teoría-Avances. IISA - 8716. Santiago, Chile.
292 p.
GASTO, J.,D.PANARIO y F. COSIO. 1993. Sistema
de caracterización de ecorregiones y determinación de sitio y
condición. Red de Pastizales Andinos, Santiago, Chile. 254 p.
GRIME, J. P. 1979. Plant strategies and vegetation
processes. John Wiley and Sons, Chischester, Great Britain. 222 p.
GRIME, J. P., J. G. HODGSON y R. HUNT. 1989.
Comparative plant ecology: A functional approach to common British species.
London, Great Britain. 742 p.
MONTALDO, P. 1975. Determinación
de las zonas agroecológicas entre la Cuesta de Lastarría y el
Seno de Reloncaví, Chile. Agro Sur (Chile) 3(2): 117-130.
MONTALDO, P. y R. PESSOT. 1974. Cambios sinecológicos
en una pradera bajo " influencia de talajeo, fertilización, quema
y competencia interespecífica. Turrialba 24 (3): 265-273.
PINOCHET, D. 1990. Fertilización de
praderas permanentes de la zona Sur. In.: L. Latrille, ed. Avances en Producción
Animal. Universidad Austral de Chile, Facultad de Ciencias Agrarias, Instituto
de Producción Animal. Serie B-14. Valdivia, Chile. Pp. 181-209.
QUINTANILLA, V. 1974. La representación
cartográfica preliminar de la vegetación chilena. Ediciones de
la Universidad Católica de Valparaíso. Editorial Universitaria.
Valparaíso. 74 p.
RAMIREZ, C, C. SAN MARTIN y D. CONTRERAS.
1990. Ecosociología de los marismas litorales del Centro Sur de Chile.
Agro Sur (Chile) 18: 104-112.
RAMIREZ, C., C. SAN MARTIN, A. ELLIES y V.
FINOT. 1991. El valor indicador ecológico de las malezas del Centro Sur
de Chile. Agro Sur (Chile) 19: 94-116.
RODRIGUEZ, J. 1993. Manual de fertilización.
Editorial Alfabeta Impresores. Santiago, Chile. 362 p.
SIERRA, C. 1991. Características de
acidez de seis suelos de la Xa Región y respuesta del trigo al encalado.
INIA, Estación Experimental Remehue. Boletín Técnico (Chile)
N° 179. 14 p.