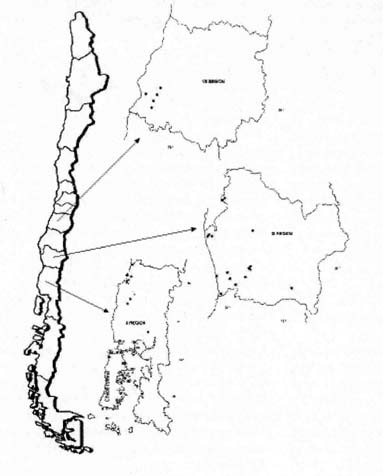
Murta (Ugri molinae T), germplasm collect sites, regions VII, IX and X region.
Ivette Seguel B., Enrique Peñaloza
H., Nelba Gaete C. Adolfo
Montenegro B. y Andrea Torres P.
Instituto de Investigaciones Agropecuarias
Centro Regional de Investigación Carillanca.
Casilla 58-D. Temuco Chile.
Recepción de originales: 15 de
Enero 1999
1 Financiado por FONDECYT 1960032
COLLECTING AND MOLECULAR CHARACTERISATION MURTA (UGNl MOLINAE TURCZ.) GERMPLASM IN CHILE.
Key words: Ugni molinae, germplasm collecting, soil chemical
characterisation. molecular characterisation.
One hundred accessions of murta (Ugni molinae Turez,} were collected
during the 1996 season in an area extending from the VII to the X Región
(35° to 42º SL). Sites of collection were characterised by marine climatic
and soils with low levels of N and available P, low levels of exchangeable K
and high levels of Al saturation. Fruits shown an extensive variability ín
chemical and physical properties with both size and weight decreasing from North
to South and soluble solids increasing . A clear relationship between euvironmental
conditions and the expression of fruit colour was found, with pink and yellow
fruits being more frequent towards the North whereas red fruits were common
towards the coldest southern area. Variability within the population was confirmed
at molecular level by applyíng the random amplified polymorphic DNA (RAPD)
technique. By using 31 polymorphic loci generated by seven primer, similarities
among accessions ranged from 0.100 a 0.833 and from 0.182 to 0.909 when Jaccard's
and Nei and Li's coefficient of similarity where computed. These results show
and extensive genetic variation within the population of Ugni molinae
T. evaluated, and suggest the feasibility to irnprove commercial attributes
of this species by single plant selection.
Palabras claves: Ugni molinae, colección de gemoplasma, caracterización
del suelo, característica molecular.
Se colectaron 100 accesiones de murta (Ugni molinae Turez) durante la temporada
1996 en un área que se extiende desde la Vll a la X Región {35°
a 42°LS). Los sitios de colecta se caracterizaron por clima marino y suelos
con bajos niveles de. N y P disponible, K intercambiable y altos niveles de
saturación de A 1. Los frutas presentaron una variabilidad significativa
respecto a características físico-químicas, se determinó
que el peso y el tamaño de los frutos incrementan de Norte a Sur a diferencia
de los sólidos solubles, cuyo incremento se observa de Sur a Norte. Se
determinó una clara relación entre las condiciones ambientales
y la expresión del color de los frutos. Mientras los colores amarillo
y rosado fueron más frecuentes hacia el Norte, los frutos rojos fueron
más comunes en el área Sur. La variabilidad dentro de la población
se confirmó mediante un análisis molecular a través de
RAPD's. Utilizando 31 locus polimorficos generados por siete partidores, se
determinó que la similitud entre accesiones fluctuó entre 0.100
a 0.833 y desde 0.182 a 0.909 según el coeficiente de Jaccard y Nei&Li
respectivamente. Los resultados muestran una amplia variabilidad genética
dentro de !a población de Ugni molinae T.. lo que sugiere la posibilidad
de mejorar atributos comerciales de la especie mediante selección de
plantas individuales.
En Chile existen alrededor de 6.265 especies de plantas, de las cuales un 14,5
% son introducidas y un 85,5 % lo constituyen especies originarias de Chile,
clasificadas como nativas y endémicas (Marticorena, 1994).
Dentro de este grupo de especies se encuentra la murta o murtilla (Ugni molinae
Turcz, Myrtaceae), especie silvestre, frecuente de encontrar asociada a otras
especies que forman parte del bosque nativo del sur de Chile. La planta produce
frutos globosos y pequeños, de agradable sabor y aroma, los que son utilizados
para el consumo fresco, fabricación artesanal de mermeladas, jarabes,
postres y licores (Novoa, 1983).
La natura! adaptación de esta especie a suelos de baja fertilidad, unido
a su habilidad para competir con otras plantas, indican que U. molinae podría
constituirse en una interesante alternativa no sólo para suelos degradados,
sino también como una opción para el sector frutícola del
sur del país. En este sentido, se menciona la factibilidad de introducir
esta especie en los mercados nacionales e internacionales como fruto fresco
o procesado, ampliando su espectro actual de usos. Investigaciones han demostrado
que la semilla contiene un alto grado de ácidos grasos insaturados como
consecuencia de su elevado contenido de ácido linoleico (78.7%) y ausencia
de ácido linolénico, indicando su potencial como aceite comestible
(Pessa y Caprile, 1993).
En términos prácticos, la incorporación de Ugni molinae
a la fruticultura nacional, implica necesariamente el inicio de un programa
sistemático de investigación tendiente a solucionar las principales
limitantes de producción, de manera que permitan transformar este fruto
silvestre en un fruto cultivado. Experiencias en este sentido se han realizado
con éxito en especies de fruto pequeño, originalmente silvestres,
tales como el cranberry (Vaccinium macrocarpon), el arándano o blueberry
(Vaccinium corymbosum), el lingonberry (Vaccinium vitis-idaea)y sanddon (Hippophae
rhamnoides), cuyo mejoramiento genético ha dado origen a una importante
producción de frutos destinados al consumo fresco y la agroindustria
(Buzeta, 1997). Sobre la base del potencial que presentaría este material
endémico, el Instituto de Investigaciones Agropecuarias inició,
a través de un proyecto FONDECYT, tareas de prospección, recolección
y caracterización de Ugni molinae, con el propósito de obtener
materiales que pudieran eventualmente transformarse en un nuevo cultivo, exclusivo
en el ámbito mundial. En este trabajo se informa sobre la prospección,
colecta, y caracterización molecular de murta o murtilla (Ugni molinae
Turcz,), colectada en el sur de Chile.
Cobertura de la colecta y metodología de muestreo
El área de colecta cubrió un amplio rango de condiciones agroecológicas
de Chile, desde la Vll hasta la X regiones, en el transecto de la Cordillera
de la Costa a Cordillera de los Andes. La selección de los sitios de
colecta se basó en antecedentes bibliográficos sobre la distribución
de la especie informados en la literatura (Alba, 1977; Landrum,
1988; Hoffmann, 1994). La distancia entre los sitios de
colecta dependió de la uniformidad de los ecosistemas, seleccionándose
aquellos con características disímiles en cuanto a ubicación
geográfica y antecedentes edafoclimáticos.
La colecta de germoplasma se realizó entre febrero y mayo de 1996, considerando
la época de fructificación de la especie. El criterio utilizado
para individualizar las accesiones dentro de cada sitio se basó en características
fenotípicas observables tanto de la planta, como de los frutos, dirigiéndose
la colecta hacia aquellos que presentaron mayor divergencia. En cada sitio de
colecta se tomaron muestras de frutos maduros, ramillas de crecimiento para
las etapas de regeneración, y material de herbario para la determinación
de las asociaciones botánicas. Paralelamente se obtuvieron muestras de
suelo superficiales (0-20 cm), en cada sitio de colecta de germoplasma, para
su caracterización química. Se determinó nitrógeno
inorgánico disponible, fósforo disponible Olsen, pH en agua (relación
1/2,5), materia orgánica, cationes de intercambio (Ca, Mg, Na, K), suma
de bases, aluminio intercambiable, CIC efectiva y saturación de aluminio
según metodología de Sadzawka (1990).
Para la documentación (datos de pasaporte) de las accesiones, se utilizó
una ficha de colecta de germoplasma, con información referente a: localización
del sitio de colecta, fuente de recolección, tipo y frecuencia de la
muestra, asociaciones botánicas, descripción de suelo, y aspectos
fenotípicos de la planta. Los datos de origen de las accesiones fueron
georeferenciados mediante un GPS (Geographic Positional System) y digitalizados
en mapas a través de un Sistemada Información Geográfica
(SIG).
Caracterización físico-química de los frutos.
Con el propósito de obtener información sobre algunas características
morfológicas y químicas "in situ", se colectaron y analizaron
frutos de murta, evaluándose color, peso, diámetro ecuatorial
(promedio de 20 frutos por accesión), y contenido de sólidos solubles.
El color de los frutos se determinó utilizando como referencia el atlas
de colores de Kuppers (1996), en tanto que los sólidos
solubles se obtuvieron mediante refractometría (expresado en ºBrix),
a partir de una muestra de 20 frutos por accesión.
Caracterización molecular de las accesiones
La caracterización morfológica de las accesiones se complementó
con estudios de variabilidad genética realizados utilizando la técnica
del ADN polimórfico amplificado al azar (RAPD), basado en la reacción
de la polimerasa en cadena (PCR). El análisis molecular se realizó
en 45 accesiones de murta representativas de todos los sitios de colecta. (Cuadro
1, figura 1). En total se evaluaron 33 partidores seleccionados al azar,
considerando sólo aquellos con alto contenido de Guanina-Citosina (60
y 70%). Del total de partidores inicialmente evaluados, se seleccionaron siete
para los estudios posteriores, sobre la base de la generación de productos
amplificados reproducibles. El ADN se extrajo desde hojas en crecimiento según
procedimiento de Doyle y Doyle (1987), con algunas modificaciones.
El tejido (0,1 g peso fresco) se maceró en nitrógeno líquido,
se homogenizó con 0,7 ml de tampón de extracción (100 mM
Tris HCI; pH 8.0; 20 mM EDTA; 1,4 M NaCI; 2% CTAB; 2 % PVP y 0,3% ß-mercatoethanol)
y se incubó a 65°C x 30 min. Posteriormente, se adicionó dos
volúmenes de cloroformo-isoamyl alcohol (24:1) y se centrifugó
a 10.000 rpm x 5 min. Los ácidos nucleicos se precipitaron con dos volúmenes
de 95% de etanol y se peletizaron porcentrifugación a 5.000 rpmx3 min.
El "pellet" se lavó dos veces en 80 y 70 % de etanol y se resuspendió
en 100 µL de tampón TE. Los extractos se incubaron con RNAsa A
(Sigma) a 37°C x 1 h y se almacenaron a 4°C hasta su utilización.
|
|
| Figura 1. Sitios de colecta de germoplasma de murta (Ugri molinae
T), VII, IX, X Región - Chile Murta (Ugri molinae T), germplasm collect sites, regions VII, IX and X region. |
La reacción de la polimerasa en cadena (PCR) se realizó en un
volumen total de 16 mL conteniendo tampón 1XPCR (Gibco BRL), 1,5 mM MgCI2,
0,1 mM de cada deoxinucleotido (dATP, dCTP, dGTP y dTTP), 0,2 mM partidor (Operon),
1,0 unidad Taq DNA polymerasa (Biorad), y 4 mL de DNA genómico. El protocolo
para RAPD consistió en un paso inicial de denaturación de 2 min
a 94°C, seguido de 40 ciclos a 94°C x 25 seg, 35°C x 1 min, y 72°C
x 1,5 min, en un termociclador Perkin Elmer 480. Los fragmentos amplificados
se separaron por electroforesis en geles de agarosa al 1,4% (p/v) conteniendo
bromuro de etidio. visualizados en un transiluminador y fotografiados con película
Polaroid 665. Todos los análisis se realizaron con repeticiones
La posición de las bandas generadas por RAPD se transformó en
una matriz de caracteres binarios (1 para presencia y 0 para ausencia de una
banda en una posición particular), realizándose la comparación
entre bandas mediante análisis multivariado (NTSYS-pc, versión
2,02). El grado de heterogeneidad entre accesiones se determinó utilizando
los coeficientes de similitud de Jaccard y de Nei y Li, de acuerdo al programa
NTSYS-pc, versión 2,02.
Distribución geográfica y habitat de crecimiento
Se colectaron 100 accesiones de Ugni molinae Turcz, en 36 sitios. (Cuadro 1;
Figura 1). La determinación botánica de las accesiones colectadas
permitió concluir que el 93% corresponde a Ugni molinae Turcz. y un 7,0%
a Ugni candollei Barn, especie que se encontró principalmente asociada
a sectores costeros de la IX y X regiones.
La amplia distribución latitudinal de la especie, observada en la colecta
de germoplasma realizada entre la Vll y X regiones, sugiere que la murta o murtilla
se desarrolla en diversas condiciones climáticas. Longitudinalmente,
la mayor frecuencia de la especie, se determinó desde la depresión
intermedia (72° LE) hacia la Cordillera de la Costa (74° LE). En un
gradiente altitudinal, las accesiones colectadas se obtuvieron entre 10 y 250
msnm (Figura 1). En relación al clima, según Novoa
y Villaseca (1989), en la Vll Región se colectó en sectores
donde el clima característico corresponde a mediterráneo marino,
en tanto que en la IX y X regiones, los materiales se colectaron en climas de
tipo marino húmedo patagónico, marino cálido, marino frío
y marino fresco, dependiendo de la ubicación de los sitios de colecta.
En su habitat natural, la especie se encontró preferentemente en terrenos
no cultivados, a orillas de caminos, en bosques naturales o en manchones ("murtales"),
alejado de centros urbanos y, por lo tanto, de baja presión antrópica.
Se determinó que U. molinae crece asociada a especies nativas, con un
alto predominio de Pernettya mucronata, Luma apiculata, Aristotelia chilensis,
Embothrium coccineumy Rhaphitamnus spinosus (Cuadro 2). En
general, las especies asociadas a la murta, se desarrollan en terrenos húmedos,
cercanos a riberas de ríos y lagos, conformando los bosques del Centro
Sur de Chile (Hoffmann, 1994).
| Cuadro 2. Especies asociadas a Ugni molinae
Turcz., en diferentes sitios de colecta. Species associated with Ugni molinae Turcz. across differentcollection sites. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
El análisis químico de las muestras de suelo obtenidas en los
36 sitios de colecta y, tomando como referencia el diagnóstico preliminar
de los niveles de fósforo, nitrógeno, potasio, materia orgánica
y pH de los suelos de la IX Región (Montenegro, 1991),
se concluyó que un 97,1 % de los sitios presentaron suelos con niveles
inferiores a 5 ppm de fósforo disponible (rango considerado muy bajo),
que un 76,4 % mostró niveles de nitrógeno disponible inferiores
a 9 ppm (muy bajos) y que un 61,7% presentó niveles de potasio intercambiable
inferiores a 100 ppm (muy bajo a bajo).
En el 50% de los sitios, los suelos presentaron niveles de materia orgánica
(MO) considerados bajos y muy bajos, en tanto que, en el 82 % de los sitios
prospectados los suelos se ubicaron en un rango de pH que varió entre
5,6 - 6,0. Respecto de la saturación de aluminio, el 59,2% de los sitios
presentaron suelos con niveles superiores a 15,1%, considerado, según
Gallardo y Borie (1999) y Borie et al. (1999),
muy alto y detrimental para varios cultivos tales como cebada, trigo y raps.
De acuerdo a la información, el ambiente natural de U. Molinae, se caracterizó
por suelos bajos en fósforo y nitrógeno disponible, bajos en potasio
intercambiable, y con altos porcentajes de saturación de aluminio (Cuadro
3). Estos antecedentes sugerirían que U. molinae sería una
especie que se adapta a condiciones de estrés nutricional, aún
cuando no se dispone de antecedentes que permitan predecir su comportamiento
en suelos sin limitantes químicas, con fertilización, o bajo manejo
agronómico. En las condiciones de suelo antes señaladas, la especie
en estudio se desarrolla formando plantas arbustivas de altura variable, entre
0.3 m y 1,70 m, y excepcionalmente, puede alcanzar una altura superior a los
3m (sitio 26, Putique-Chiloé).
| Cuadro 3. Distribución de la población
de suelos de los sitios de colecta en rangos correspondientes a diferentes
parámetros químicos del suelo. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Características de los frutos colectados.
Al analizar las características de los frutos en las diferentes accesiones
se observó que el diámetro ecuatorial fluctuó entre 0,71
y 1,31 cm con un promedio de 0,94 cm, consistente con lo indicado en la literatura
(Landrum, 1988). Es importante señalar que la variación
latitudinal afectó significativamente este carácter, de manera
que mientras el 56% de las accesiones colectadas en la X Región presentaron
frutos con diámetro ecuatorial superior a 1,0 cm, sólo el 14%
de los frutos alcanzó este tamaño en la Vll Región. Al
respecto, Lavín y Vega (1996) reportan que el diámetro
ecuatorial de la murta colectada en la Vll Región fluctúa entre
0,5 y 0,9 cm, lo que concuerda con el presente análisis, en donde e!
50% de los frutos colectados en esta región poseen diámetros ecuatoriales
entre 0,71 y 0,89 cm (Figura 2).
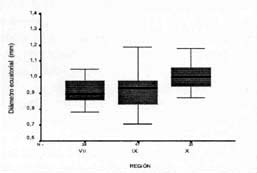 |
| Figura 2. Diámetro ecuatorial de frutos de murta (Ugni
molinae T.), colectados en las regiones VII, IX, X. Chile. Ecuatorial diameter value of murta (Ugni molinae T.) fruit collected in regions VII, IX and X. Chile. |
El peso de los frutos del material colectado fluctuó entre 0,21 y 1,01 g, presentando un rango más amplio que lo reportado por Hevia et al. (1993) y Landrum y Donoso (1990), quienes señalaron variaciones entre 0,42 y 0,51 g. De manera similar a la tendencia que caracterizó al diámetro ecuatorial, el peso de los frutos aumentó en un gradiente latitudinal norte-sur. De esta manera, en la Vll Región no se encontraron accesiones con frutos superiores a 0,61 g, en tanto que, en las regiones IXy X, se apreció que un 4%y 20% de las accesiones, respectivamente, presentaron frutos con pesos superiores a este valor (Figura 3).
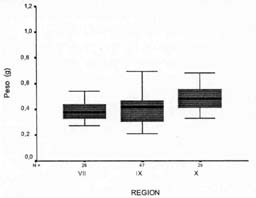 |
| Figura 3. Peso (g) de frutos de murta (Ugni molinae T.),
colectados en las regiones VII, IX, X. Chile. Weigth value of murta (Ugni molinae T.), fruit collected in regions VII, IX and X. Chile. |
Los sólidos solubles presentaron un amplio rango de dispersión
que fluctúo entre, 6,5 y 28° Brix (Figura 4). En promedio, los sólidos
solubles de frutos provenientes de la Vll, IX y X regiones correspondieron a
18,16; 16,88 y 13,58° Brix, respectivamente. En la Vll Región la
mayoría de las accesiones (86%) se ubicó entre 14 y 21.5°
Brix, en la IX Región el 59 % de ellas se ubicó en este rango
y en la X Región sólo el 48%. Sobre la base de estos antecedentes,
los sólidos solubles del fruto de murta disminuyen de norte a sur, una
tendencia opuesta a lo que ocurre con el diámetro ecuatorial y el peso.
En la X Región no se detectaron frutos con sólidos solubles superiores
a 22° Brix, a diferencia de lo observado en las regiones Vll y IX donde
un 11% y 17% y de las accesiones, respectivamente, presentaron frutos con parámetros
superiores a este valor, considerado alto si se compara con el promedio de 15º
Brix obtenido en evaluaciones realizadas en la IX Región en arándano
alto (Vaccinium corymbosum) (Fígueroa, 1991).
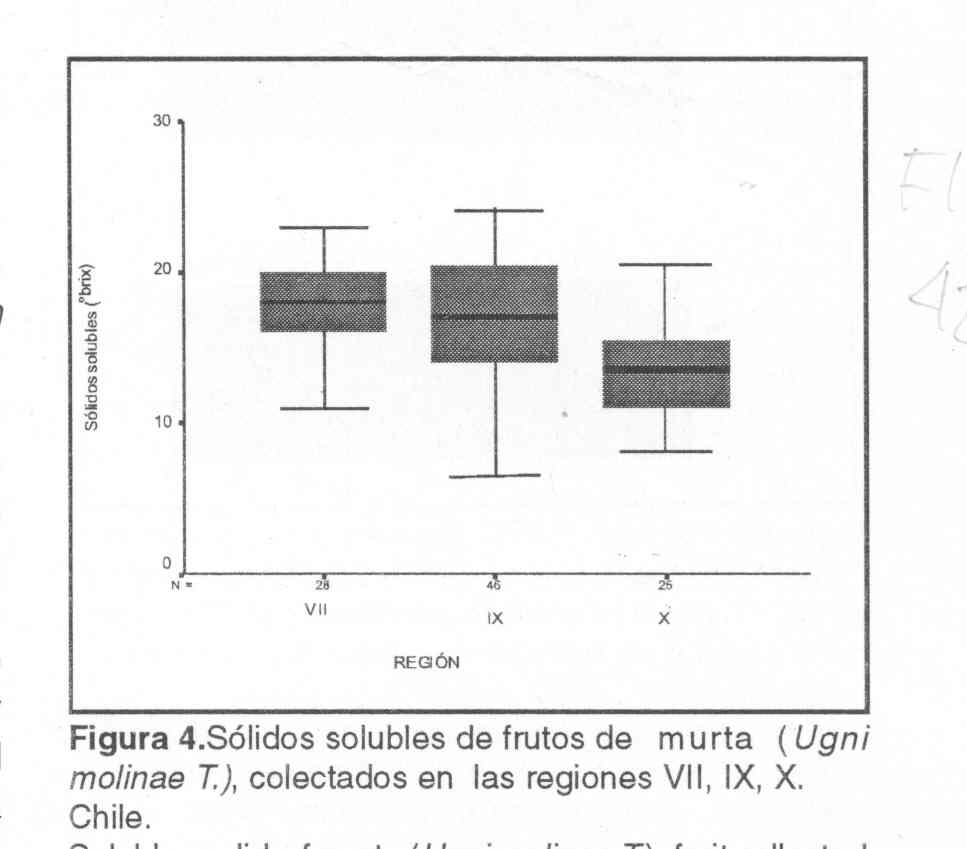 |
| Figura 4. Sólidos solubles de frutos de murta (Ugni molinae
T.), colectados en las regiones VII, IX, X. Chile. Solubles solid of murta (Ugni molinae T.), fruit collected in regions VII, IX and X. Chile. |
De lo anterior se desprende que existe una relación latitudinal en
cuanto a los caracteres del fruto: diámetro ecuatorial, peso y sólidos
solubles. El diámetro ecuatorial y peso de los frutos aumenta de norte
a sur del país, situación inversa a lo observado para los contenidos
de sólidos solubles. La accesión de mayor peso y diámetro
ecuatorial proviene de la localidad Cutipay en la Isla de Chiloé, X Región,
y la accesión con mayor contenido de sólidos solubles (28º
Brix) proviene de la localidad Piren Alto, VII Región.
Se determinó una gran variabilidad en el color de los frutos, observándose
tonalidades desde blancas a rojo oscuro, con una tendencia a una mayor expresión
del color rojo en las zonas más frías. Considerando un gradiente
latitudinal de norte a sur, se determinó que en los frutos de murta de
la Vll Región predominan los tonos rosado amarillentos, en la IX Región
predominan los frutos de color rojo, aún cuando un número importante
de accesiones es de color rosado, y en la X Región destacan los frutos
de color rojo intenso.
La variación fenotípica observada en un gradiente latitudinal
para los caracteres: peso, diámetro ecuatorial, sólidos solubles
y color del fruto, podría explicarse en términos de variación
clinal, entendiéndose por esta, al cambio gradual de un carácter,
en especies de amplia distribución geográfica, como resultado
de la adaptación a un cambio continúo del medio ambiente (Langlet,
1963; Grant, 1971). Las principales fuentes de variación
clinal, en extensas áreas de distribución de las especies, son
los factores climáticos (Stern & Roche, citados por Donoso,
1995). Esto sugiere la posibilidad de explotar la variación clinal
mediante el movimiento de genotipos entre zonas climáticas de modo de
permitir la expresión de un carácter deseado, mientras no revierta
a su fenotipo original.
Caracterización molecular
Del total de partidores seleccionados (7 de un total de 33 inicialmente evaluados,
OPAN-19 y OPAP-03 se destacaron por presentar el mayor polimorfismo. Treinta
y una bandas fácilmente identificables se analizaron por la presencia
o ausencia de fragmentos de ADN amplificado, los que fluctuaron entre 600 y
2000 pares de bases. Perfiles representativos generados por tres de los siete
partidores utilizados en este estudio(OPAN 19, OPAP 03y OPAK02} se presentan
en la Figura 5. El grado de relación entre accesiones de U. molinae,
determinado mediante la derivación de coeficientes de similitud, no varió
significantemente cuando se utilizaron los coeficientes de Jaccard o Nei y Li,
con una correlación entre matrices de 0,99, determinada de acuerdo al
Test de Mantel (NTSYS-pc, versión 2,02). De acuerdo a estos resultados,
no se observó homología entre las 45 accesiones colectadas en
un amplio rango de ambientes del sur del país, con coeficientes de similitud
que fluctuaron entre 0,1 y 0,833, y entre 0,182 y 0,909 para Jaccard y Nei y
Li, respectivamente. La ausencia de homología se observó incluso
entre accesiones colectadas en un mismo sitio geográfico (Tapihue), como
se muestra para la serie MUR-06colectada en la VII Región (Cuadro 1,
Figura 1).
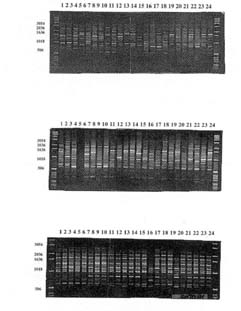 |
| Figura 5. Patrones RAPD generados con partidores OPAN-19 (panel
superior) OPAP-03 (panel central) y OPAK-17 (panel inferior). El orden de
carga de izquierda a derecha es marcador de peso molecular-accesión
1-24. RAPD patterns generated with primers OPAN-19 (upper, OPAP-03 (middle) and OPAK-17 (lower). From left to right, weigth standar, accession 1 to 24. |
Aun cuando la significativa variabilidad genotípica entre accesiones
es consistente con lo observado en caracteres morfológicos y químicos,
es importante mencionar que el procedimiento de colecta, dirigido hacia la selección
de genotipos divergentes, podría sobrestimar el valor real de divergencia
dentro de la población. El amplio rango de variabilidad, sin embargo,
permite señalar la posibilidad cierta de mejorar los atributos comerciales
de esta especie mediante selección de plantas. Se ha sugerido que la
heterogeneidad morfológica de U. molínae podría ser el
resultado de su aparentemente fácil polinización cruzada (Wilckens
et al. 1991), confirmado por los estudios de Seguel et al,
(1999), quienes demostraron un 47 % de polinización cruzada para
esta especie, en donde el genero Bombus spp es el principal agente polinizador.
Consecuentemente, la heterogeneidad observada en la población analizada
podría estar asociada al hábito reproductivo de la especie, o
al efecto del ambiente sobre el comportamiento de su principal agente polinizador.
Si esto último es verdadero, entonces la mayor divergencia genética
debería encontrarse en accesiones colectadas en áreas o ambientes
que favorezcan la polinización entomófila, como lo observado en
este trabajo para regiones más boreales.
Debido al valor potencial observado en la especie Ugni molinae, se consideró
necesario mantener un banco de germoplasma con el propósito de disponer
de material para futuras evaluaciones. Actualmente, el 100% de la colección
es conservada en INIA Carillanca forma vegetativa en jardines de campo. Paralelamente,
se están desarrollando investigaciones tendientes a establecer protocolos
para la conservación in vitro de la especie.
DONOSO, Z,C. 1995. Bosques Templados de Chile y Argentina. Variación, Estructura y Dinámica. Ecología Forestal. Editorial Universitaria. Santiago. Chile. 485 p.
DOYLE, J. J. Y DOYLE, J.L 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19: 11-19.
FIGUEROA, G.E, 1991. Efecto de tres modalidades de raleo en seis variedades de arándano alto (Vaccinium corymbosum L.) Tesis. Ingeniero Agrónomo, Universidad de la Frontera. Facultad de Ciencias Agropecuarias. Temuco. Chile, 85 p.
GALLARDO, F. Y BORIE, F 1999. Sensibilidad y tolerancia de especies y cultivares a condiciones de acidez. Test rápido de diagnóstico. Frontera Agrícola. Temuco, Chile. 5(1-2):3-18.
BORIE, F.; GALLARDO, F; MORA, M.; GARCÍA, 1999. Sensibilidad y tolerancia a la acidez de los cultivos en condiciones de campo, Frontera Agrícola. Temuco. Chile. 5(1-2):19-28.
GRANT, V. 1971, PlantSpeciation. Columbia University Press. New York. U,S.A,284 p.
HEVIA, H. F.; VENEGAS, V. A.; WILCKENS, E. R.; ARAYA, S. .F.; TAPIA, V. 1993. Murtilla (Ugni molinae T.) III. Algunas características del fruto colectado en Chile. Agro-Ciencia. Santiago. Chile. 9 (1):63-66.
HOFFMAN, J. A. 1994. Flora silvestre de Chile, Zona Araucana. Fundación. Claudio Gay. Santiago.Chile. 258 p.
KUPPERS, H. 1996. Atlas de colores. Editorial Blume.Barcelona. España ,161 p.
LANDRUM, L.R. 1988. The myrtle family (Myrtaceae) in Chile. Proceeding of the California Academy of Science.47 (12): 277-317.
LANDRUM, L.R. Y DONOSO, C. 1990. Notes on Economic Plants. Economic Botany. 44 (4): 536 -539.
LANGLET, 0.1963. Patterns and terms of intra-specific Ecological variability. Nature, 200. (4904).
LAVÍN.A.A. Y VEGA, M. A. 1996. Caracterización de frutos de murtilla (Ugni molinae turcz.) en el área de Cauquenes. Simiente. Santiago. Chile. 64 (3): 93.
MARTICORENA, C. 1994. Contribución a la estadística de la flora vascular de Chile. Gayana Botánica, (Chile) 47 (3-4): 85-113.
MONTENEGRO, B.A. 1991. Diagnóstico preliminar de los tenores de nitrógeno, fósforo, potasio, materia orgánica y pH de los suelos de la IX Región. Investigación y Progreso Agropecuario Carillanca. Temuco. Chile. 10(3);3-11.
NOVOA , H. R, 1983. Antecedentes sobre establecimiento y producción de frutos de la especie Ugni molinae Turcz. Tesis Ingeniero Forestal. Universidad Austral de Chile. Valdivia, Chile. 90 p.
NOVOA, R. Y VILLASECA, S. 1989. Mapa agroclimático de Chile. Instituto de Investigaciones Agropecuarias. Santiago. Chile, 221 p.
PESSA, J.A. Y CAPRILE, L. R. 1993,Análisis ycomposición de aceite en semilla de murta(Ugni molinae). Alimentos. Santiago. Chile 18(1): 11-14.
SADZAWKA, R. A. 1990. Métodos de análisis de suelos. Instituto de Investigaciones Agropecuarias. Serie La Platina Nº 16. Santiago. Chile. 130 p.
SEGUEL, I; RIVEROS, M., LEHNEBACH, C. Y TORRES, A. 1999. Antecedentes reproductivos y fenológicos de Ugni molinae Turcz. ( Myrtaceae). Phyton. 65:13-21
WILCKENS, R.; VENEGAS, A.; HEVIA, F.;TAPIA, M.; ARAYA,F. Y CONEJEROS, S. 1991 Murtilla (Ugni molinae T.) I. Obtención de extractos foliares no oxidados para separar electroforeticamente proteínas y enzimas en geles de poliacrilamida. Agro Ciencia 7 (2): 129-134.