CARACTERISTICAS DEL ALMIDON Y CONTENIDO DE PROTEINA
DE QUINOA (CHENOPODIUM QUINOA W.) CULTIVADA BAJO DIFERENTES
NIVELES DE NITROGENO EN CHILLAN1
Felícitas Hevia H.2,
Rosemarie Wilckens E.3 Marisol Berti D.3
y Ricardo Badilla B.
2 Facultad de Ing. Agrícola, Universidad de Concepción,
Casilla:537 Chillán, Chile, e-mail: fhevia@udec.cl.
3 Facultad de Agronomía, Universidad de Concepción,
Casilla: 53 7 Chillán, Chile
Recepción de originales:20 de Noviembre
del 2000
ABSTRACT
STARCH CHARACTERISTICS AND PROTEIN CONTENT OF QUINOA (CHENOPODIUM
QUINOA W.) GROWN UNDER DIFFERENT NITROGEN LEVELS IN CHILLAN
Key words: Chenopodium quinoa, Starch, Protein, Differential
Scanning Calorimetry.
The effect of the nitrogen fertilization was evaluated on some physico-chemical
and thermic properties of starches, protein content and protein yield ha-1
of the genotypes of quinoa Faro and UDEC10. Sodium nitrate was applied in doses
of 0; 75; 150 and 225 N kg ha-1. The average protein content varied
between a 13,5 to 15,0%. It was higher for the application 225 N kg ha-1.
The average protein yield increased from 134,1 to 408,3 kg ha-1.
The degree of gelatinization, the water absorption index and the swelling power
were not affected by nitrogen fertilization, while starch solubility was affected.
Their average values, respectively were the following 10,3%; 2,7g; 2,9g and
7,2%. The granules of perisperm starch were organized in compact aggregates,
differences among them were not observed.
RESUMEN
Palabras claves : Chenopodium quinoa, Almidón, Proteína,
Calorimetría diferencial de barrido.
Se evaluó el efecto de la fertilización nitrogenada sobre algunas
propiedades físico-químicas y térmicas del almidón,
contenido del proteína y rendimiento de proteína ha-1
de los genotipos de quinoa Faro y UDEC10. Se aplicó salitre sódico
en dosis de 0; 75; 150 y 225 kg N ha-1. El contenido promedio de
proteína varió entre un 13,5 a 15,0%. Sólo fue superior
con la aplicación de 225 kg N ha-1. El rendimiento promedio
de proteína aumentó desde 134 a 408 kg ha-1. El grado
de gelatinización, el índice de absorción de agua y el
poder de hinchamiento no fueron afectados por la fertilización.
INTRODUCCION
La quinoa (Chenopodium quinoa W.) es un pseudocereal que fue usado como
un alimento básico por los antiguos pueblos de Sudamérica. Actualmente
se cultiva en Argentina, Bolivia, Chile, Colombia, Ecuador y Perú (Wahli,
1990). En países como Alemania se estudia la posibilidad de introducirlo
en la dieta diaria (Aufhammer et al., 1995) y en EEUU
en usar su almidón en la confección de polietilenos biodegradables
(Alexander, 1996).
La semilla de quinoa se usa en la preparación de platos a la forma entera
y partida, harina cruda y tostada, hojuelas y polvo instantáneo. Se ha
observado que en la preparación de pan, galletas y tortas cantidades
superiores al 15 o 20% de harina de quinoa, le otorgan un sabor desagradable
(Ruales y Nair, 1992). El contenido de proteína de
la semilla varía, según el cultivar, entre 10 y 22% y su valor
biológico es similar al de la caseína (Ruales y
Nair, 1992; Wahli, 1990).
Las características del almidón juegan un rol fundamental en
el desarrollo de nuevos productos alimenticios, debido a la gama de propiedades
funcionales de éste como espesante (sopas), formador de gel (gomas),
estabilizador coloidal (salsas) y otras (Pérez, 1996).
Entre las pruebas que se realizan se encuentran el grado de gelatinización,
el termograma, el índice de absorción, el poder de hinchamiento
y la solubilidad en agua. Estas características del almidón influyen
en la palatabilidad de los alimentos preparados (Ruales et
al., 1993). Sin embargo, no existen antecedentes en los cuales se les
relacione con el manejo del cultivo.
El objetivo de este estudio fue evaluar el efecto de la dosis de nitrógeno
sobre las características físico-químicas y térmicas
del almidón y el contenido de proteína de dos genotipos de quinoa
sembrados en Chillán.
MATERIALES Y METODOS
Semilla de quinoa. Se utilizaron semillas de quinoa 'Faro' y 'UDEC10',
sembradas en septiembre de 1995 en la Estación Experimental de la Facultad
de Agronomía de la Universidad de Concepción, Campus Chillán.
Las semillas fueron recolectadas en la VIII Región, 'Faro' en el Fundo
El Faro, comuna El Carmen y 'UDEC10' en la zona costera. La siembra se realizó
en un suelo de la serie Diguillín de textura franca, plano, de buen drenaje
y antes del ensayo presentaba 7,8 mg kg-1 N-NO3; 25,1
mg kg-1 P; 6,17% de materia orgánica.
Para asegurar la disponibilidad de fósforo y potasio en el cultivo se
aplicó, al voleo, 100 unidades de P2O5 como superfosfato
triple y 50 unidades de K2O como sulfato de potasio por ha. El fertilizante
nitrogenado utilizado fue salitre sódico y correspondió a 0, 75,
150 y 225 kg N ha-1, en dos parcialidades, la mitad cuando presentaba
cuatro hojas verdaderas y el resto a inicio de panoja.
Eliminación de las saponinas. Se procedió de acuerdo al
método descrito por Junge y colaboradores (1975).
Cocción de la semilla. En 200 mL de agua destilada se calentó
a ebullición durante 30 minutos 30 g de semilla de quinoa lavada, y luego
se dejó secar durante 12 h en una estufa Binder con aire forzado a 55°C.
Molienda de la semilla cruda y cocida. Se realizó en un molino
Retsch modelo ZM-2, con un tamiz de 250 mesh.
Materia seca. Se determinó en 2,500 g de quinoa molida cruda
y cocida, en una estufa Binder con aire forzado a 105°C durante 12 h.
Características físico-químicas del almidón.
Indice de absorción de agua (IAA), poder de hinchamiento (PH) e índice
de solubilidad en agua (ISA). Se determinaron de acuerdo al método descrito
por Hevia et al. (1999).
Microscopía electrónica de barrido. Se usó semilla
cruda sin lavar de cada tratamiento y se procedió según la metodología
descrita por Hevia y colaboradores (1999), usando un microscopio
electrónico de barrido Siemens Autoscan U-l del Laboratorio de Microscopía
Electrónica de la Universidad de Concepción.
Calorimetría diferencial de barrido. Los termogramas se realizaron
en el Departamento de Polímeros de la Facultad de Ciencias Químicas
de la Universidad de Concepción. Se utilizó semilla molida de
quinoa lavada cruda y cocida de ambos genotipos, con tres niveles 0, 75 y 150
kg N ha-1. En una suspensión acuosa (60% p/v), las harinas
fueron calentadas hasta 240°C a una velocidad de 10°C min-1
(Ruales y Nair, 1992). Del termograma se obtuvo la entalpia (VH) y las temperaturas
de gelatinización (de inicio, máxima y de término de gelatinización,
To, Tp y Te, respectivamente). A partir de los DH
de la muestra cruda y cocida se determinó el grado de gelatinización
del almidón (GGA), por medio de la ecuación (%)=[1-(DHcocida/DHcruda)]
100 (Holm et al, 1988).
Proteína. Se determinó por el método Kjeldahl en
0,250 g de quinoa molida cruda (AOAC, 1997). El resultado se
informó como porcentaje, base materia seca, usando el factor 6,25.
Rendimiento de proteína por hectárea. Se determinó
a partir del porcentaje de proteína y el rendimiento en semilla (comunicación
personal, C.A. Méndez, 1997).
Diseño experimental. Se utilizó los diseños de
bloques completos al azar con arreglos de parcela dividida y subdividida, en
que los factores en estudio fueron genotipo (parcela principal), dosis de nitrógeno
(subparcela) y cocción (subsubparcela).
En parcela dividida para las características físico-químicas
del almidón y de proteína se usaron cuatro dosis de N con cuatro
repeticiones, y para las características térmicas de la gelatinización
se utilizaron tres dosis de N con tres repeticiones. En cambio, en parcela subdividida
se usaron las mismas dosis de N, pero con tres repeticiones.
Análisis estadístico. Previa transformación de
los datos porcentuales mediante la relación (x+0,5)1/2 (Little
y Jackson, 1978), se realizó los análisis de varianza de acuerdo
a los diseños utilizados. Para medir las diferencias entre medias de
tratamientos se utilizó el test de diferencias mínimas significativas
(DMS) con un 95% de confianza.
RESULTADOS Y DISCUSION
Características físico-químicas del almidón.
Para el índice de absorción de agua (IAA) y poder de hinchamiento
(PH) de la semilla cruda molida no se encontró diferencia entre genotipo
ni entre los cuatro niveles de N aplicado (Cuadro 1). Esto
indica que el genotipo y la fertilización nitrogenada no habrían
influido sobre la capacidad del almidón para absorber y retener agua
(Ruales et al., 1993).
| Cuadro 1: Características físico-químicas
del almidón de semilla molida cruda y cocida de dos genotipos de
quinoa, fertilizados con cuatro dosis de nitrógeno.
Table 1: Starch physico-chemical characteristics of raw and cooked
quinoa seed of two genotypes fertilized with four nitrogen rates.
|
|
Dosis de N
(Kg ha-1)
Promedio |
Crudo |
|
Cocido |
|
|
| Faro
|
UDEC10
|
Promedio
|
Faro
|
UDEC10
|
|
|
Indice de absorción de agua (IAA, g)
|
| |
| 0 |
2,679 |
2,732 |
2,705 A |
4,541 bc(2) |
4,596 bc |
4,569 |
| 75 |
2,666 |
2,622 |
2,644 A |
4,477 bc |
5,826 a |
5,151 |
| 150 |
2,733 |
2,647 |
2,689 A |
4,542 bc |
4,416 c |
4,479 |
| 225 |
2,791 |
2,656 |
2,723 A |
4,528 bc |
4,699 b |
4,614 |
| Promedio |
2,717 A 1 |
2,664 A |
|
4,522 |
4,884 |
|
| |
|
|
|
|
|
|
| Poder de hinchamiento
(PH, g) |
| |
| 0 |
2,899 |
2,943 |
2,921 A |
4,787 b |
4,794 b |
4,791 |
| 75 |
2,901 |
2,816 |
2,858 A |
4,725 b |
6,049 a |
5,387 |
| 150 |
2,957 |
2,836 |
2,896 A |
4,759 b |
4,558 c |
4,658 |
| 225 |
3,039 |
2,838 |
2,938 A |
4,629 c |
4,321 d |
4,476 |
| Promedio |
2,949 A |
2,858 A |
|
4,725 |
4,931 |
|
| |
|
|
|
|
|
|
| Indice de solubilidad
en agua (ISA, %) |
| |
| 0 |
7,560 b2 |
7,145 bc |
7,352 |
5,143 |
4,123 |
4,633 A |
| 75 |
7,509 b |
6,302 d |
6,905 |
4,940 |
3,039 |
3,989 B |
| 150 |
7,525 b |
6,662 cd |
7,093 |
4,547 |
3,170 |
3,859 B |
| 225 |
8,349 a |
6,572 cd |
7,460 |
4,567 |
3,502 |
4,034 B |
| Promedio |
7,736 |
6,672 |
|
4,799 A |
3,458 B |
|
|
|
|
|
|
1 letras mayúsculas
distintas en sentido vertical y horizontal indican diferencia entre los
efectos principales, según prueba DMS (P £
0,05).
2 Letras minúsculas diferentes en sentido vertical y
horizontal indican diferencias entre la interacción genotipo x
fertilización, según prueba DMS (P £
0,05).
|
Respecto al índice de solubilidad en agua (ISA) se observó interacción
genotipo x fertilización, fue mayor en 'Faro' cuando se aplicó
225 kg N ha-1, en cambio el de 'UDEC10' fue menor con 75 kg N ha-1 , correspondiendo
al mayor y menor grado de degradación del almidón y por ende,
de modificación del mismo (Ruales et al, 1993).
En la semilla cocida se presentó interacción genotipo x fertilización
(P £ 0,001),
tanto para el IAA como para el PH, en donde 'UDEC10' fertilizado con 75 kg N
ha-1 fue superior a los restantes tratamientos (Cuadro 1). En el
ISA se observó diferencias, tanto entre genotipo como entre fertilizaciones,
siendo superior en 'Faro' y cuando no se aplicó N. Los resultados de
'UDEC10' fertilizado con 75 kg N ha-1 dejan entre ver que un mayor
IAA se relaciona con un mayor PH y un menor ISA, al igual como lo señala
Puga (1991) para una mezcla de arroz-frejol.
Para medir el efecto de la cocción sobre las características
físico-químicas del almidón se utilizaron los resultados
de las semillas crudas y cocidas. Se presentó interacción genotipo
x fertilización x cocción (Cuadro 2), tanto
para el IAA (P £ 0,05)
como para el PH (P £ 0,01),
siendo los valores promedios mayores en las semillas cocidas y entre éstas,
para las de 'UDEC10' fertilizadas con 75 kg N ha-1.
Esto concuerda con lo señalado por otros autores. Así, Ruales
y Nair (1994) informaron que al someter la semilla a cocción se destruyen
los agregados de almidón y se dañan los granulos, lo que facilitaría
la absorción y retención del agua por parte de éstos.
Según Fennema (1980), al incrementar la temperatura
las moléculas de almidón vibran vigorosamente, rompiéndose
los enlaces intermoleculares, y separándose fragmentos de la cadena de
almidón, facilitando la formación de uniones puente hidrógeno
con el agua. Por otra parte, el calentamiento del almidón destruiría
el complejo lípido-amilosa, que interfiere con la absorción del
agua. Por otro lado, los enlaces puente H que se forman entre la amilosa y el
agua, son también responsables de la absorción de agua y de la
gelatinización del almidón.
Para el ISA se observó diferencias (P £ 0,001),
tanto entre genotipo como por la cocción, siendo mayores los valores
promedios para 'Faro' y para las semillas crudas (Cuadro 2).
Por efecto de la cocción el ISA promedio de ambos genotipos disminuyó
en un 42%. Esto se debería, a que al subir la temperatura por sobre los
80°C, tanto amilasas como amilopectinas se solubilizan en el sobrenadante
(Fennema, 1980). Pero también se pudieron solubulizar
proteínas, vitaminas y minerales, durante la cocción (Meiner,
1978). Lo anterior explicaría el menor ISA observado en la semilla
molida cocida.
Los valores promedios obtenidos para el IAA y pH de las semillas crudas fueron
superiores a los informados previamente por Ruales et al
(1993) y por Hevia et al. (1999). Mientras que
para las semillas cocidas el IAA fue similar y el pH inferior a los informados
por Ruales et al. (1993). En cambio, el ISA en ambos
casos fue ligeramente inferior.
De acuerdo a los antecedentes informados por Puga (1991),
las semillas de 'UDEC10' fertilizadas con 75 kg N ha-1 serían
muy apropiadas para la elaboración de snacks, bebidas y papillas de alta
concentración para lactantes, que requieren de productos de alto grado
de cocción y de un mayor IAA. En cambio, si se quiere preparar papillas
instantáneas, sopas o colados, que necesitan de productos de un alto
IAA y de un mayor ISA, las más indicadas serían las de 'Faro'
sin fertilizar.
| Cuadro 2: Indice de absorción de agua y poder de hinchamiento
de semillas molidas crudas y cocidas de dos genotipos de quinoa para la
interacción genotipo x cocción e índice de solubilidad
en agua, con relación a genotipo y cocción.
Table 2: Water absorption index and swelling power of milled quinoa
seeds of two genotypes, row and cooked; and the interaction among genotypex
Nitrogen rate and cooking treatment and genotipyxcooking interaction for
water solubility.
|
|
| Genotipo |
Kg N ha-1 |
Crudo |
Cocido |
Promedio |
|
| Indice de absorción
de agua (g, IAA) |
| |
|
|
|
|
| Faro |
0 |
2,679 c(1) |
4,541 b |
3,610 |
| UDEC10 |
0 |
2,732 c |
4,596 b |
3,664 |
| Faro |
75 |
2,666 c |
4,477 b |
3,572 |
| UDEC10 |
75 |
2,622 c |
5,826 a |
4,224 |
| Faro |
150 |
2,733 c |
4,542 b |
3,638 |
| UDEC10 |
150 |
2,647 c |
4,416 b |
3,532 |
| Faro |
225 |
2,791 c |
4,528 b |
3,660 |
| UDEC10 |
225 |
2,656 c |
4,699 b |
3,678 |
| Promedio |
|
2,690 |
4,703 |
|
| |
|
|
|
|
| Poder de hinchamiento
(g, PH) |
| |
| Faro |
0 |
2,899 c |
4,787 b |
3,843 |
| UDEC10 |
0 |
2,943 c |
4,794 b |
3,869 |
| Faro |
75 |
2,901 c |
4,725 b |
3,813 |
| UDEC10 |
75 |
2,816 c |
6,049 a |
4,433 |
| Faro |
150 |
2,957 c |
4,759 b |
3,858 |
| UDEC10 |
150 |
2,836 c |
4,558 b |
3,697 |
| Faro |
225 |
3,039 c |
4,629 b |
3,334 |
| UDEC10 |
225 |
2,838 c |
4,321 b |
3,579 |
| Promedio |
|
2,690 |
4,703 |
|
| |
|
|
|
|
| Indice de solubilidad (%,
ISA) |
| |
|
|
4,799 |
6,268 A |
| Faro |
|
7,736 |
3,458 |
5,065 B |
| UDEC10 |
|
6,672 |
4,144 B |
|
| Promedio |
|
7,171 A(2) |
|
|
|
|
|
|
| |
1Letras minúsculas diferentes en sentido
vertical y horizontal indican diferencias entre las interacciones triples,
según prueba DMS (P £
0,05).
2Letras mayúsculas distintas en
sentido vertical y horizontal indican diferencias entre los efectos principales,
segun prueba DMS (P £
0,05). |
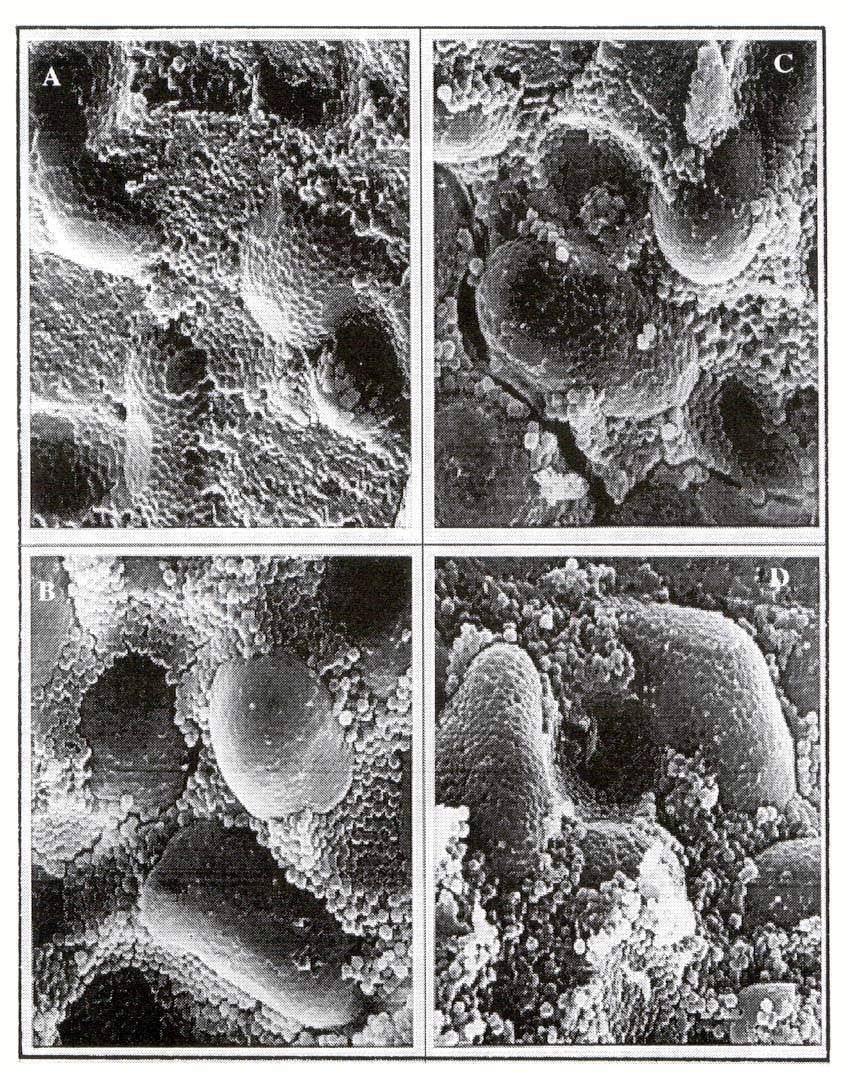
Microestructura del granulo de almidón. En las microfotografías
del perisperma de las semillas de quinoa (Figura 1) se puede
observar que los gránulos de almidón son de forma esférica
a poligonal y de pequeño tamaño. Estos se presentan aislados o
conformando agregados alargados, esféricos a oblongos, que se encuentran
rodeados por una gran cantidad de granulos más simples. Una estructura
similar describieron Ruales y Nair (1994) y Hevia
et al. (1999). En general, no se observó diferencia, en la
estructura y distribución de los granulos de almidón, entre los
tratamientos.
|
|
| |
|
Figura 1: Microfotografías del perisperma de quinoa "Faro"
sembrada en Chillán bajo distintas dosis de nitrógeno, A)
0 kg N ha-1; B 75 kg N ha-1; C) 150 kg N ha-1
y D) 225 kg N ha-1.
Figure 1: Microphotographs of quinoa perisperm "Faro" sown
in Chillán with different nitrogen rates. A) 0 kg N ha-1:
B) 75 kg N ha-1; C) 150 kg N ha-1 y D) 225 kg N
ha-1.
|
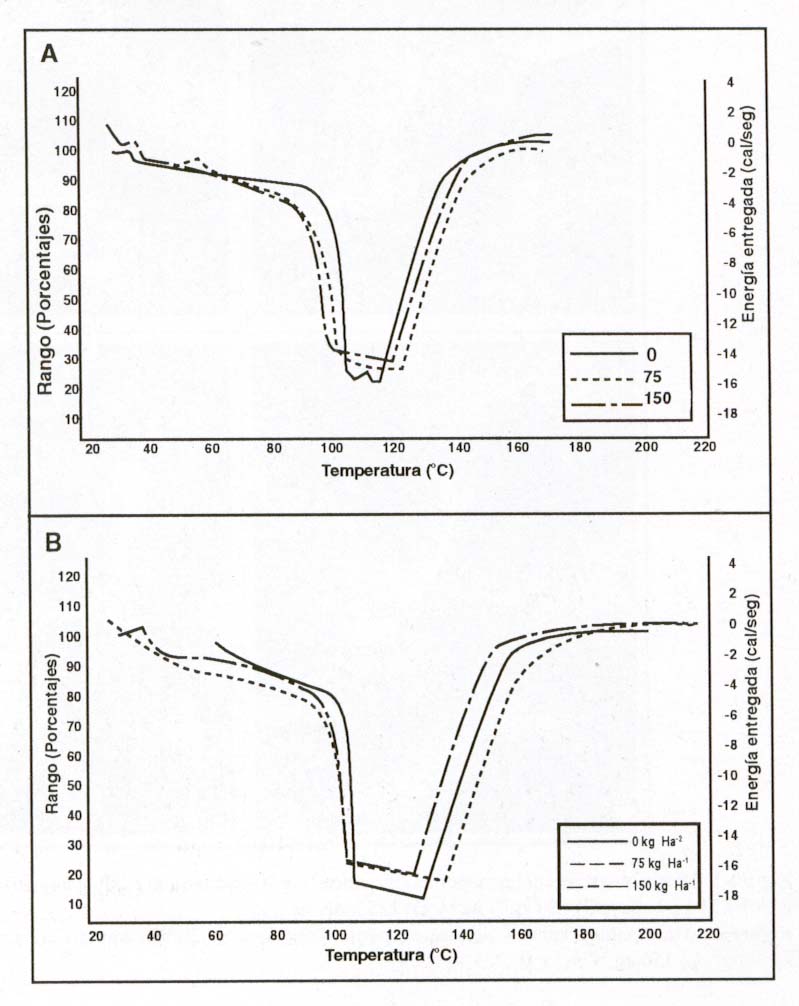
Características térmicas de la gelatinización del almidón.
Los valores de las temperaturas de gelatinización (de inicio, máxima
y de término de gelatinización, To, Tp y Te, respectivamente)
y de la entalpia (DH)
de la misma, fueron obtenidos a partir de los termogramas de la semilla de quinoa
molida cruda (Figura 2) y cocida. Dado el alto costo del análisis
sólo se consideraron tres niveles de nitrógeno, dejando fuera
las muestras correspondiente a la mayor dosis.
|
|
| |
|
Figura 2: Termogramas de las harinas crudas de las semillas de
dos genotipos de quinoas A) Faro y B) UDEC10, sembradas en Chillán
bajo distintas dosis de nitrógeno (0, 75 y 150 kg ha-1).
Figure 2: Raw milled seed thermograms from of two quinoa genotypes
A) Faro and B) UDEC10 with different nitrogen rates.
|
En las semillas molidas crudas (Cuadro3) se observó
diferencia entre genotipo (P £
0,05) para la Te, la que fue superior en 'UDEC10'.Además, se observó
diferencias entre las dosis de N para To, Tp, Te y H.
Todas las variables fueron menor en los tratamientos fertilizados con 75 kg
N ha-1 e igual en aquellos con 0 y 150 kg N ha-1. Estos resultados fueron superior
a los informados previamente por Wahli (1990), Ruales
et al. (1993) y Ruales y Nair (1994).
Según Fennema (1980) la gelatinización y las
características de los geles formados dependen no sólo de la temperatura,
sino que más bien de los tipos y cantidades de otros compuestos que acompañan
al almidón en el perisperma, como son los azúcares, proteínas,
lípidos, ácidos y agua. Estos pueden retardar o incluso llegar
a inhibir la gelatinización. En consecuencia, la dosis de N, a través
de estos compuestos presentes en las harinas, sería capaz de alterar
las características térmicas de la gelatinización.
Por otra parte, en las semillas molidas cocidas (Cuadro 3)
se observó diferencias (P £
0,05) entre las dosis de N para DH,
siendo inferior el valor promedio de ambos genotipos en aquellas fertilizadas
con 75 kg N ha-1.
| Cuadro 3: Características térmicas de la gelatinización
de semilla molida de dos genotipos de quinoa molidas crudas y cocidas,
en relación a la fertilización nitrogenada.
Table 3: Gelatinization characteristics of milled quinoa seeds,
raw and cooked, regarding nitrogen rates
.
|
|
Dosis de N
(Kg ha-1)
|
|
Cruda |
|
|
Cocida |
|
|
|
|
Faro |
UDEC10 |
Promedio |
Faro |
UDEC10 |
Promedio |
|
|
|
Temperaturas de inicio de gelatinización
(Tº, ºC)
|
|
|
|
0
|
100,59
|
100,66
|
100,63 A
|
97,77
|
99,16
|
98,46 A
|
|
75
|
72,33
|
89,19
|
80,76 B
|
96,69
|
98,32
|
97,51 A
|
|
150
|
96,31
|
99,27
|
97,79 A
|
96,46
|
96,14
|
96,29 A
|
|
Promedio
|
89,74 A (1)
|
98,03 A
|
|
96,97 A
|
97,87 A
|
|
|
|
|
|
|
|
|
|
|
Temperatura de máxima gelatinización
(Tp, ºC)
|
|
|
|
0
|
106,35
|
103,92
|
105,14 A
|
103,71
|
104,56
|
104,14 A
|
|
75
|
91,81
|
103,59
|
97,70 B
|
103,42
|
103,79
|
103,61 A
|
|
150
|
102,27
|
103,48
|
102,88 A
|
100,15
|
103,65
|
101,90 A
|
| Promedio |
100,14 A
|
103,66 A
|
|
102,64 A
|
104,00 A
|
|
| |
|
|
|
|
|
|
|
Temperatura final de gelatinización
(Te, ºC)
|
|
|
| 0 |
117,37
|
129,19
|
123,28 A
|
154,11
|
144,84
|
149,47 A
|
| 75 |
100,36
|
124,21
|
112,29 B
|
141,31
|
137,18
|
139,24 A
|
| 150 |
120,01
|
128,89
|
124,55 A
|
142,20
|
139,11
|
140,65 A
|
| Promedio |
112,58
|
127,43 A
|
|
145,87 A
|
140,37 A
|
|
| |
|
|
|
|
|
|
|
Entalpía de gelatinización
(DH,
cal/g)
|
| |
| 0 |
267,73
|
265,66
|
266,69 A
|
218,20
|
239,73
|
228,96 A
|
| 75 |
193,71
|
230,51
|
212,11 B
|
170,48
|
209,58
|
190,03 B
|
| 150 |
239,68
|
253,34
|
246,51 A
|
224,65
|
236,54
|
230,59 A
|
|
Promedio
|
233,71 A
|
249,83 A
|
|
204,87 A
|
228,61 A
|
|
|
|
|
| Letras mayúsculas distintas en sentido vertical
y horizontal indican diferencia, según prueba DMS (£
0,05) |
Para determinar si hubo un efecto de la cocción sobre estas variables,
se utilizaron los resultados obtenidos con las semillas molidas crudas y cocidas.
Se presentó interacción genotipo x cocción para Te (P £ 0,05).
Las semillas crudas presentaron el menor valor, y entre éstas las de
'Faro' (Cuadro 4).
Se observó diferencias en el DH
(P £ 0,05),
entre genotipo debido a la cocción, siendo mayor el valor promedio para
'UDEC10' y cuando no hubo cocción (Cuadro 4). Se presentó,
además, interacción fertilización x cocción, en
que las semillas molidas cocidas fertilizadas con 75 kg N ha-1 presentaron el
valor promedio más bajo.
Esta última diferencia obedecería a que los tratamientos cocidos
requieren de menos energía calórica que los crudos para su gelatinización,
ya que tanto los enlaces lipido-amilosa como los amilosa-amilopectina se habrían
destruido durante la cocción. Así, se habrían formado cadenas
de menor peso molecular. Estas se unirían más fácilmente
al agua, aumentando la absorción y retención de la misma, facilitando
la gelatinización en la semilla molida cocida.
Para el grado de gelatinización del almidón (GGA) no se observó
diferencia entre genotipos ni entre las dosis de N (Cuadro 1).
Esto indicaría que el genotipo y la fertilización con N no habrían
influido sobre esta variable. Se encontró una correlación significativa
e inversa entre dosis de N y GGA (r = -0,694). Por otro lado, los resultados
obtenidos fueron inferiores a los informados por Ruales et
al. (1993) y Ruales y Nair (1994).
| Cuadro 4: Temperatura final de gelatinización de semillas
molidas crudas y cocidas de dos genotipos de quinoa para la interacción
genotipo x cocción; y entalpía de gelatinización
de acuerdo al genotipo y la fertilización, con interacción
fertilización x cocción.
Table 4: Interaction effects (genotype x cooking) for gelatinization
conclusion temperature, and enthalpy, for milled, raw and cooked quinoa
seeds, regarding with genotypes and nitrogen levels.
|
|
Genotipo/
kg N ha-1 |
Crudo |
Cocido |
Promedio |
|
| Temperatura final de gelatinización
(ºC, Te) |
| |
|
|
|
| Faro |
112,58 c(1) |
148,08 a |
130,33 |
| UDEC10 |
127,43 b |
145,98 a |
136,70 |
| Promedio |
120,01 |
147,03 |
|
| |
|
|
|
| Entalpía de gelatinización
(cal/g, DH)
|
| |
|
|
|
| Faro |
233,71 |
204,87 |
219,29 B |
| UDEC10 |
249,83 |
228,61 |
243,22 A |
| Promedio |
241,77 A (2) |
216,74 B |
|
| |
|
|
|
| 0 |
266,69 a |
233,12 b |
249,90 |
| 75 |
212,11 c |
190,03 d |
201,07 |
| 150 |
246,39 abQ |
227,09 bc |
236,74 |
| Promedio |
241,65 |
226,75 |
|
|
|
|
1Letras minúsculas diferentes en sentido
vertical y horizontal indican diferencias entre las interacciones, según
prueba DMS (P £
0,05).
2Letras mayúsculas distintas en
sentido vertical y horizontal indican diferencias entre los efectos principales,
segun prueba DMS (P £
0,05). |
Contenido de proteína. El contenido promedio de proteína
de los tratamientos fluctuó entre 12,17 y 15,16% (Cuadro
5), coincidiendo con los rangos informados por otros autores (Whali,
1990; Ruales y Nair, 1992; .
Sólo se observó diferencias entre dosis de N (P £
0,001), en la medida que aumentó la fertilización. Así,
el contenido promedio de proteína de ambos genotipos fue inferior con
150 kg N ha-1 y superior con 225 kg N ha-1. Los resultados confirman lo señalado
por Avila y Etchevers (1988), quienes indicaron que los requerimientos
nutricionales de la quinoa parecieran ser más altos que los de otras
especies cultivadas también en Chillán.
En este sentido, Herrera (1986) en avena y Hevia
et al. (1987) en trigo, observaron que una dosis de N baja a moderada incrementa
la producción de grano, pero disminuye o mantiene el contenido proteico,
en comparación con el testigo no fertilizado. Sin embargo, cuando fertilizaron
con niveles mayores de N obtuvieron incrementos lineales y significativos en
el contenido proteico de la semilla. Esto se debería a que en un principio
el cultivo, dispuso de suficiente N para un mayor desarrollo vegetativo y una
mayor formación de semilla con respecto al sin fertilizar. Luego, durante
el período de llenado de grano se habría generado una demanda
de N tan grande que ni el aporte del suelo ni la removilización desde
los tejidos de sostén, fueron suficiente para satisfacerla. Como consecuencia,
se produjo una dilución de la proteína en la semilla del material
fertilizado con 150 kg N ha-1 (Covacevich, 1987).
Es posible que este mismo fenómeno fisiológico sea el responsable
de las diferencias observadas en la solubilidad y en las características
térmicas de la gelatinización. Bajos niveles de N en el sistema
de la planta contribuyen a una menor tasa de asimilación neta de C02,
puesto que el N es el constituyente vital de la clorofila y de las enzimas de
carboxilación (Meinzer y Zhu, 1998).
El contenido promedio de proteína (Cuadro 5) para
la dosis de 150 kg N ha-1 (13,0%) fue inferior a los informados por Etchevers
(1980) y Pino (1998) y superior a los registrados por Hevia
et al. (1995) y Fuentes (1996) para la misma dosis
de N, localidad y genotipo. Estas diferencias podrían atribuirse a las
condiciones edafoclimáticas de la localidad (Hevia et
al.
(1999).
| Cuadro 5: Grado de gelatinización del almidón,
contenido de proteína y rendimiento de proteína ha-1,
para dos genotipos de quinoa cultivados bajo diferentes fertilizaciones
nitrogenadas e interacción genotipo x fertilización.
Table 5: Genotype x nitrogen interaction for degree of gelatinization,
protein content and protein yield of milled quinoa seeds for two genotypes
of quinoa.
|
|
| |
Fertilización nitrogrenada
(kg N ha-1)
|
|
| |
0
|
75
|
150
|
225
|
Promedio
|
|
|
Grado de gelatinización del
almidón (GGA, %)
|
| |
|
|
|
|
|
| Faro |
15,39
|
11,99
|
9,20
|
-
|
12,19 A
|
| UDEC10 |
9,76
|
8,90
|
6,63
|
-
|
8,43 A
|
| Promedio |
12,58 A (1)
|
10,45 A
|
7,92 A
|
-
|
|
| |
|
|
|
|
|
|
Contenido de proteína (%)
|
| |
|
|
|
|
|
| Faro |
13,45
|
13,06
|
12,17
|
14,85
|
13,38 A
|
| UDEC10 |
13,69
|
13,27
|
13,85
|
15,00
|
13,99 A
|
| Promedio |
13,57 B
|
13 16 BC
|
13,01 C
|
15,00 A
|
|
| |
|
|
|
|
|
|
Rendimiento de preteína (kg
ha-1)
|
| |
|
|
|
|
|
| Faro |
134,92 e(2)
|
229,32 d
|
244,89 dc
|
309,83 cb
|
229,74
|
| UDEC10 |
133,32 e
|
287,65 db
|
355,37 b
|
506,63 a
|
320,74
|
| Promedio |
134,12
|
258,49
|
300,13
|
408,30
|
|
| |
|
|
|
|
|
|
|
|
|
1 Letras mayúsculas distintas
en sentido vertical y horizontal indican diferencia entre los efectos principales,
según prueba DMS (P £
0,05).
2 Letras minúsculas diferentes en sentido vertical y horizontal
indican diferencias entre la interacción genotipo x fertilización,
según prueba DMS (P £
0,05) |
Rendimiento de proteína por hectárea. Se observó
interacción genotipo x fertilización (P £ 0,01).
En la medida que se aumentó el aporte de N, el rendimiento de proteína
ha-1 fue mayor para ambos genotipos (Cuadros 5). Sin embargo, 'UDEC10' presentó
una mayor respuesta a la fertilización nitrogenada.
Los mayores rendimientos de proteina no necesariamente coincidieron con los
mayores contenidos de proteína. Hevia et al. (1994)
en trigo, observaron que el rendimiento de semilla influye en mayor medida que
el contenido de proteína sobre esta variable.
Los rendimientos de proteína de 'Faro' y 'UDEC10' fueron menores a los
informados por Fuentes (1996) y Pino (1998). Esto se debería
a que el rendimiento de semilla es influenciado por las condiciones ambientales
de la temporada durante el desarrollo del cultivo (Pino, 1998).
CONCLUSIONES
En base a los resultados obtenidos se pudo concluir que la dosis de fertilizante
nitrogenado no afectó el índice de absorción de agua, el
poder de hinchamiento ni el grado de gelatinización. Pero sí influyó
sobre el índice de solubilidad en agua, la entalpia de gelatinización,
el contenido de proteína y el rendimiento de proteína por hectárea.
AGRADECIMIENTOS
Memoria del cuarto autor para optar al título de Ingeniero Agrónomo.
Proyecto N°96.122.104-1 Dirección de Investigación Universidad
de Concepción.
BIBLIOGRAFIA
ALEXANDER, R. 1996. Starch in plastics. Cereal Foods World
41:426-42.
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. 1997. oficial
methods of analysis. 11ª ed. Horwitz (Ed.). Washington, D.C., U.S.A.
AUFHAMMER, Y, J. LEE, E. KÜBLER, M. KUHN und S. WAGNER.
1995. Anbau und Nützung der Pseudocerealien Buchweizen (Fagopyrum esculentum
Moench), Reismelde (Chenopodium quinoa W.) und Amaranth (Amaranthus
ssp. L.) ais Kornerfruchtarten. J. für landwirtschaftliche Forschung
46(2): 125-126.
AVILA, P. Y J. ETCHEVERS. 1988. Efecto de la profundidad de
siembra y la fertilización NPK sobre la emergencia, producción
de materia seca y composición química de la quinoa {Chenopodium
quinoa W.). Agro-Ciencia 4(1):5-10.
COVACEVICH, C. 1987. Dosis y época de aplicación
de nitrógeno sobre la calidad en un trigo de primavera cv. Onda. Agro-Ciencia
3(2):117-123.
ETCHEVERS, G. 1980. Composición química de algunas
leguminosas y chenopodiaceas de la provincia de Ñuble. Cienc. Inves.
Agrar. 7:191-196.
FENNEMA, O. 1980. Química de los alimentos. Editorial
Acribia ,S.A. Zaragoza. España.
FUENTES, X. 1996. Contenido de proteínas y saponinas
de 29 genotipos de quinoa (Chenopodium quinoa W.) de distinta procedencia
cultivados en Chillán. Tesis Ing. Agrónomo. Univ. Concepción,
Chillán, Chile.
HERRERA, B. 1986. Caracterización fisiológica
de la Avena sativa L. cv. Nehuen en relación a la nutrición
nitrogenada. Tesis Ing. Agrónomo. Univ. Concepción, Chillán,
Chile.
HEVIA, F, J. FUENZALIDA y C. COVACEVICH. 1987. Influencia
de la fertilización nitrogenada sobre la calidad panadera y el contenido
de proteína del trigo de primavera cv. Onda. I. Dosis de fertilizante
nitrogenado. Agro-Ciencia 3(2):117-123.
HEVIA, F, R. WILCKENS, J. FUENZALIDA y T. VIDAL. 1994. Estructura
del endosperma y patrones electroforéticos de proteínas de tres
cultivares de cebada sembrados en tres localidades en Chile. Agro-Ciencia 10(2):85-92.
HEVIA, F., R. WILCKENS, M, URBINA, L. FINOT and M. BERTI.
1995. Quinoa seeds biopolymers. III. Symposium on new crops, new opportunities,
new technologies. Indianapolis, U.S.A. 22-25 Octubre.
HEVIA, F.,M. BERTI, R. WILCKENS, HUMBERTO SERRI y A. PINO.
1999. Contenido de proteínas, saponinas y algunas características
del almidón en semillas de quinoa sembradas en diferentes fechas y localidades.
Agro-Ciencia 14(2):293-302.
HOLM, J., I. LUNDQUIST, I. BJÔRCK and A. ELIASSO. 1988.
Degree of starch gelatinization, digestion rate of starch in vivo, and
metabolic response in rats. Am. J. Clin. Nutri. 47: 1010-1016.
JUNGE, I., P. CERDA y K. ALID. 1975. Lupino y quínoa;
Estado actual de los conocimientos y de las investigaciones sobre su empleo
en la alimentación humana. Universidad de Concepción. Escuela
de Ingenería Civil. Concepción, Chile.
LITTLE, T. y F. JACKSON. 1978. Métodos estadísticos
para la investigación en la agricultura. Ed. Trillas. México.
MEINER, C. 1978. Proximate composition and yield of raw and
cooked mature dry legumes. J. Agric. Food Chem. 24(6):1122-1125.
MEINZER, F. and J. ZHU. 1998. Nitrogen stress reduces the
efficiency of the C4CO2 concentrating system, and therefore
quantum yield, in saccharum (sugarcane) species. Journal Experimental Botany
49(324): 1227-1234.
PÉREZ, E. 1996. Algunas experiencias sobre la modificación
química del almidón. Instituto de Ciencias y Tecnología
de Alimentos. Facultad de Ciencias, Caracas, Venezuela.
PUGA, E. 1991. Estudio de algunas características funcionales
y organolépticas de mezclas arroz-frejol precocidas por extrusión
utilizables en alimentos infantiles. Escuela Politécnica Nacional, Facultad
de Ing. Química, Quito, Ecuador.
RUALES, J. and B. NAIR. 1992. Nutritional quality of the protein
in quinoa (Chenopodium quinoa W.) seeds. Plant Foods Human Nutr. 42:1
-11.
RUALES, J., S. VALENCIA and B. NAIR. 1993.Effect of processing
on the physico-chemical characteristics of quinoa flour (Chenopodium quinoa
W.). Starch/Stärke 45:13-19.
RUALES, J. and B. NAIR. 1994. Properties of starch and dietary
fibre in raw and processed quinoa (Chenopodium quinoa, Willd.) seeds.
Plant Foods Human Nutr. 45:223-246.
WAHLI, C. 1990. La Quinoa: hacia su cultivo comercial. Latinreco
S.A. Quito, Ecuador.