*Cesar Núñez F., *Juan J. Cantero G.,
*Mirian L. Petryna L., *María A. Amuchastegui B., **Carlos Ramírez G**.
* Universidad Nacional de Río Cuarto, Departamento de Biología Agrícola, Facultad
de Agronomía y Veterinaria, Ruta Nacional N 36, KM 601. 5800 Río Cuarto, Córdoba,
Argentina
**Universidad Austral de Chile, Facultad de Ciencias, Instituto de Botánica,
Casilla 567, Valdivia, Chile
Recepción de originales 17 de diciembre de 2001
Effects of fire and grazing on the functional types of Poaceae C3 and C4 in a Central Argentinean mountain prairie.
Key words: Prairie, fire, grazing, Poaceae, functional types.
The effects of fire and grazing on the functional types of Poaceae C3 and C4 in Central Argentinean mountain prairies were evaluated. The study site was located between the south latitudes 32º 42' and 32º 51' and the longitudes 64º 56' and 64º 48', at an altitude of 1400 m above sea level. Two communities were studied, Festuca hieronymi (tall prairie) and Stipa juncoides-Sorghastrum pellitum (short prairie) with the following four treatments: 1) without fire, with grazing 2) without fire or grazing, 3) with fire and grazing and 4) with fire, without grazing. Fire increased the variety of C4 species and the combination of fire and grazing had a greater effect on the Poaceae C3 species in the tall prairie, while in the short prairie the absence of grazing increased the variety of Poaceae C4 species. Fire, alone, did not improve the amount of grass in the C4 component of the community in the tall prairie, but the selective grazing of the Poaceae C3 narrowed the C3/C4 relationship. A period of time without grazing in the short prairie permitted the reestablishment of the cover and diversity of the Poaceae C4 and the recovery of its productivity. These field experiments demonstrate that the combined use of fire and grazing must be used with caution, considering its different effects on each prairie community. The system of light continuous grazing can not be applied to two different communities in the same manner. In tall prairie, grazing following a fire modified the attributes of the prairie, whereas this did not occur in the short prairie.
Palabras claves: Praderas, fuego, pastoreo, Poaceae, tipos funcionales.
Se evaluaron los efectos del fuego y el pastoreo sobre los tipos funcionales de Poaceae C3 y C4, en pastizales serranos centroargentinos. El área de estudio se ubica entre los 32º 42´ y 32º 51´ de latitud Sur y los 64º 56´ y 64º 48´ de longitud Oeste, con altitud de 1400 m.s.m. Se estudiaron dos comunidades Festuca hieronymi (pastizal alto) y Stipa juncoides-Sorghastrum pellitum (pastizal bajo). Se plantearon 4 tratamientos:1) sin fuego con pastoreo, 2) sin fuego sin pastoreo, 3) con fuego con pastoreo,4) con fuego sin pastoreo. El fuego incrementó la riqueza de las especies C4, y la combinación de fuego y pastoreo afectó más a las Poaceae C3 en el pastizal alto, en cambio en el pastizal bajo la ausencia de pastoreo potenció la riqueza de las Poaceae C4. El fuego, por sí solo, no aumentó las especies C4 de la comunidad en los pastizales altos, pero el pastoreo selectivo de las Poaceae C3 estrechó la relación C3/C4. El rezago del pastizal bajo permitió reestablecer la cobertura y diversidad de las Poaceae C4 y la recuperación de su productividad. Este experimento de campo demuestra que la combinación fuego y pastoreo debe utilizarse con precaución, considerando sus efectos diferenciales sobre cada comunidad. Un sistema de pastoreo liviano y continuo, no puede ser aplicado a las dos comunidades por igual. En el pastizal alto el pastoreo postfuego ejerce efectos que modifican los atributos del pastizal, situación que no ocurre con el pastizal bajo.
En las plantas, el tipo funcional se ha definido como aquel grupo de especies
que utilizan la misma clase de recursos de un modo semejante, exhibiendo respuestas
similares a las condiciones ambientales y con efectos comunes sobre los procesos
ecosistémicos dominantes (Walker, 1992; Noble
y Gitay, 1996). Estos atributos funcionales adquieren gran importancia en
las evaluaciones de las respuestas ecosistémicas a los disturbios (Tilman
y Downing, 1994; Epstein et al., 1997; Díaz
et al., 1997; Paruelo et al., 1998). En pastizales naturales,
se ha prestado mucha atención a los tipos fotosintéticos C3 y C4,
como indicadores de cambios ambientales (Epstein et al., 1997).
Según Polley (1997) el aumento en la concentración de CO2
en la atmósfera producirá cambios notables en la composición de especies C3
y C4, en áreas de transición entre bosques y pastizales.
El estudio de las relaciones cuantitativas entre la productividad de los grupos
funcionales de Poaceae C3 y C4 y las variables ambientales
es crucial para comprender la dinámica de las comunidades vegetales de pastizales
(Epstein et al., 1997). Se ha demostrado que las especies
C4 tienen una mayor eficiencia en el uso del agua que las especies
C3 (Pearcy y Ehleringer, 1984).
El fuego y el pastoreo son los factores principales que afectan la estructura
y diversidad de las comunidades de pastizales (Sala, 1988;
Belsky, 1992; Noy-Meir, 1995; Collins,
1987) y sus efectos dependen de la historia evolutiva, de la relación planta-herbívoro
y del tipo de ecosistema (Milchunas et al., 1988). La remoción
del forraje, modifica el microclima, principalmente el espectro lumínico (Deregibus
et al., 1985) y altera las condiciones de crecimiento de las especies C3
y C4 (Steuter, 1987).
El fuego, a través de la remoción de material muerto en pié, mejora la producción
del pastizal, favoreciendo el crecimiento de las Poaceae C4 (Anderson,
1982). A su vez, el pastoreo selectivo postfuego, de las especies C3
dominantes, suele favorecer la posición competitiva de las especies C4
(Lubchencko, 1978; Ode et al., 1980). Luego
de ocurrido el evento del fuego, las Poaceae C4, generalmente responden
en forma favorable e incrementan: su vigor, la densidad de macollos y biomasa,
la floración y el mejoramiento en la producción de semillas y germinación de
las mismas (Collins y Wallace, 1990). Estas evidencias ponen
de manifiesto la importancia de las quemas al final del período de reposo invernal.
El pastoreo liviano o la supresión del fuego en pastizales altos provoca una
acumulación de material muerto y un autosombreado que podrían favorecer a las
especies C3 (Ode et al., 1980). Este material muerto
depositado sobre el suelo, puede ser un factor de control (Ketling,
1954), evitando la emergencia de plántulas y macollos de la especie dominante
(Penfound, 1964).
Steuter (1987) estudió el efecto del fuego sobre el balance
de las especies C3 y C4 en los pastizales de Norteamérica,
y los resultados obtenidos mostraron que el fuego incrementó la producción del
componente C3, independiente de la época de ocurrencia del fuego.
Este autor planteó la hipótesis de que la relación C3/C4,
en los pastizales altos, parece ser el resultado de una adaptación a largo plazo,
más que un ajuste en el corto plazo o a efectos temporales, mientras que dicha
relación en las comunidades de pastizales bajos responde a ajustes en el corto
plazo o a los cambios causados por el fuego, la humedad, la temperatura y la
luz.
Según Howe (1994), en los pastizales altos de Norteamérica,
la quema durante la estación de reposo y la exclusión al pastoreo son intervenciones
humanas que podrían promover artificialmente la dominancia de pastos altos C4
reduciendo la diversidad del pastizal. Como los períodos productivos de las
Poaceae C3 y C4 están desplazados en el tiempo (Ode
et al., 1980), el enriquecimiento del pastizal alto con especies C4
podría modificar algunos atributos del pastizal como por ejemplo, oferta forrajera
a través del año, calidad y diversidad florística, pero no existe certeza de
que ello ocurra.
En los pastizales serranos centroargentinos, aún no se ha estudiado como el
fuego, el pastoreo o su combinación, afectan las relaciones entre las Poaceae
tipo C3 y C4 en términos de diversidad y cobertura. Sólo
existen trabajos florísticos y de distribución (Cavagnaro, 1988;
Sánchez y Arriaga 1990; Cabido et al., 1997).
Bajo el punto de vista de la conservación de la diversidad vegetal y del manejo
productivo de este ecosistema, no se ha evaluado si el fuego por sí mismo o
en combinación con el pastoreo podrían ser herramientas útiles en la manipulación
de la relación de Poaceae C3/C4. Para esclarecer estos
interrogantes se plantearon los siguientes objetivos: (1) evaluar los efectos
del fuego, pastoreo y su combinación sobre la cobertura y diversidad de los
tipos funcionales C3 y C4, (2) evaluar si la situación
de manejo postfuego actual cumple con los requisitos de conservación de la diversidad
y calidad del pastizal.
1. Área de Estudio
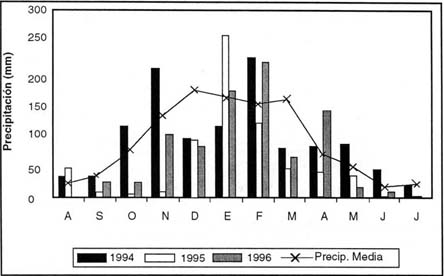
El área de estudio se ubica en el centro de Argentina entre los 32º 42´ y 32º 51´ de latitud Sur y los 64º 56´ y 64º 48´ de longitud Oeste, con una altura de 1400 m s.n.m. (Figura 1) La precipitación media anual es de 1088 mm y ella se distribuye en 2 semestres, uno lluvioso (primavera-veraño) y otro seco (otoño-invierno) (Figura 2). La temperatura media anual es de 16º C. La temperatura media máxima anual alcanza a 21ºC y la temperatura mínima media anual a 8ºC. Las heladas se inician en mayo-junio y finalizan en agosto-septiembre. El área de estudio forma parte de las Sierras de Comechingones, correspondientes a la provincia geológica de las Sierras Pampeanas. El basamento está compuesto por rocas plutónicas-metamórficas de edad Precámbrica a Paleozoica. El 60% son rocas metamórficas y un 40% rocas plutónicas, principalmente granitos (González Bonorino, 1950; Gordillo y Lencinas, 1969, Martino et al., 1995).
|
|
|
| Figura 1: |
Provincia de Cordoba, República Argentina. El área achurada indica el lugar de trabajo. |
| Figure 1: |
Province of Cordoba, Republic of Argentina. The experimental site is indicated
by the dotted area. |
|
|
|
| Figura 2: |
Régimen de precipitaciones en el sitio estudiado. |
| Figure 2: |
Rainfall pattern over the experimental site. |
Bajo el punto de vista edáfico los conocimientos son limitados y en escalas
de poco detalle. Se han reconocido grandes complejos integrados por Udorthentes
típicos, Hapludoles fluvénticos y otros Entisoles e Inceptisoles ( INTA,
1994).
La vegetación está comprendida dentro del llamado Bosque Chaqueño Serrano
(Parodi, 1964; Ragonese, 1967). Los tipos
fisonómicos dominantes son pastizales y arbustales (Kurtz, 1904;
Luti et al., 1979; Bianco et al., 1987).
Desde el punto de vista florístico es la región con mayor riqueza específica
del centro de Argentina (Cantero y Bianco, 1986) con valores
promedios de 50 especies por m2 para las diferentes comunidades vegetales
reconocidas.
2.- Tratamientos experimentales
El estudio se realizó sobre un mismo tipo litológico (gneiss) en dos unidades diferentes de paisaje, una altiplanicie ocupada por la comunidad de Festuca hieronymi (pastizal alto) y una ladera con la comunidad de Stipa juncoides- Sorghastrum pellitum (pastizal bajo). Ambas comunidades están sometidas a pastoreo contínuo con una carga animal promedio de 0.3 UA/ha (pastoreo liviano) que es la que se aplica normalmente en el pastizal. En los años 1980 y 1988 ambas situaciones fueron sometidas a un fuego prescripto. En julio de 1993 un fuego accidental afectó sólo partes de ambas unidades. En agosto de 1993 se instalaron sendas clausuras al pastoreo (900 m2) c/u., incluyendo áreas quemadas y no quemadas dentro de cada unidad de paisaje. Cuatro parcelas por unidad de paisaje/ comunidad quedaron seleccionadas. Los tratamientos considerados se indican en el Cuadro 1.
| Cuadro 1: |
Descripción de los tratamientos. | ||||||||||||||||||||||||||||||||||||||||||
| Table 1: |
Treatments description. |
||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
| *Situación de referencia (sería
la más semejante en relación a la que han evolucionado los
pastizales del centro de Argentina) ( Díaz, et al.,
1994; Cabido et al., 1997). |
|||||||||||||||||||||||||||||||||||||||||||
3. Colección y análisis de datos
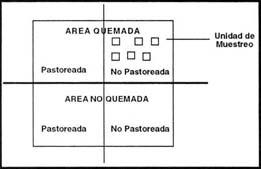
Para cada parcela del tratamiento (Figura 3), se localizaron 6 unidades de muestreo de 5 m x 5 m donde se estimó durante los veranos de 1994, 1995 y 1996, la presencia-ausencia de todas las plantas vasculares en cuadrados de 0,4 m2 distribuidos al azar, la cobertura se midió en cinco líneas de intercepción permanente de 5 m de longitud (Grey - Smith, 1964). La riqueza (S) se determinó como el número de especies presentes por unidad de superficie en cada una de las parcelas (Gastón, 1996).
 |
|
| Figura 3: |
Esquema de muestreo. |
| Figure 3: |
Experimental sampling design. |
El análisis de correspondencias libres de tendencias (DCA) ( McCune
y Mefford, 1999) fue empleado para ordenar los censos en base a su similitud
composicional. Se realizó un DCA para ordenar solamente las Poaceae C3
y C4. El reconocimiento de la vía fotosintética C4 se
realizó a través del examen de la anatomía Kranz en cortes transversales de
hojas de material fresco y de herbario y a través de la consulta bibliográfica
(Cabido et al., 1997; Sánchez y Arriaga, 1990;
Cavagnaro, 1988). Las listas de Poaceae C3 y C4
se presentan en el Anexo I. Para la nomenclatura de las especies vegetales se
siguió a Zuloaga et al., (1994). Se escogió como fecha de
muestreo el comienzo del verano, porque la mayoría de las especies se encuentran
presentes y en estado de floración y/o fructificación, lo cual permite una rápida
identificación y un muestreo más exacto de la vegetación. Este estudio se restringió
a una evaluación de los efectos de un fuego accidental y estacional sobre dos
comunidades del pastizal.
Comunidad de Festuca hieronymi
Este pastizal alto ocupa laderas escarpadas entre 1100 y 1600 m de altitud.
La especie dominante Festuca hieronymi cubre siempre más del 95% formando
grandes champas de hasta 1 m de altura. Acompañan a la especie principal Taraxacum
officinale, Margyricarpus pinnatus y varias especies del género Stipa.
Diversidad y cobertura
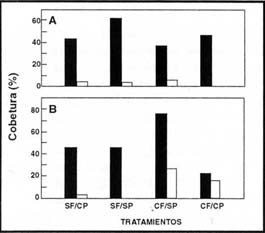
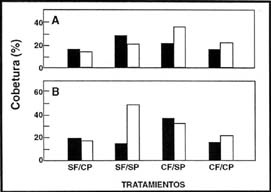
Hubo una clara asociación entre los tipos funcionales, el fuego y el pastoreo
en todos los sitios muestreados. El pastoreo luego de un año de ocurrido el
evento del fuego igualó la competencia y la cobertura de las Poaceae C3
y C4, que alcanzaron valores similares al final del ensayo (Figura
4). En este pastizal dominado por Poaceae C3, la ausencia de fuego
y de pastoreo o la ausencia de ambos, redujeron la cobertura de las Poaceae
C4, sin embargo el fuego en ausencia de pastoreo posibilitó un incremento
en su cobertura (Figura 4).
 |
|
| Figura 4: |
Cobertura de Poaceae C3 (barras negras) y C4 (barras blancas) en la comunidad de Festuca hieronimy al primer (A) y tercer (B) año de ocurrido el fuego. |
| Figure 4: |
Cover of Poaceae C3 (black bars) and Poaceae C4 (white bars) plants
in the Festuca hieronimy community during the first (A) and third
(B) years after the fires. |
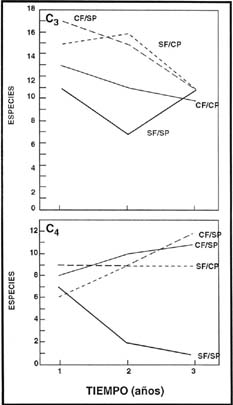
El fuego aumentó la riqueza específica y combinado con la remoción del pastoreo
se asoció a la mayor diversidad. La ausencia de fuego y pastoreo afectó negativamente
la diversidad. El fuego incrementó la diversidad de las Poaceae C4
(Figura 5).
 |
|
| Figura 5: |
Variación de la riqueza de especies de Poaceae C3 (arriba) y de Poaceae C4 (abajo) en la comunidad de Festuca hieronimy en los tres primeros años. |
| Figure 5: |
Variation of the richness of C3 Poaceae species (above) and C4 Poaceae
species (below) in the Festuca hieronimy during the first three years. |
El fuego con exclusión al pastoreo, tuvo como efecto inmediato, un incremento
en la diversidad de las Poaceae C3, que luego se diluyó, posiblemente
por efecto de la ausencia de pastoreo. La diversidad de Poaceae C4
superó levemente a las Poaceae C3. El pastoreo sin fuego (situación
de manejo actual), incrementó la diversidad de Poaceae C3 duplicando
en número a las C4, efecto que no se mantuvo a lo largo de los tres
años, ya que al finalizar el mismo, la diversidad de especies C3
y C4 fueron muy similares (Figura 5). La ausencia de fuego y pastoreo
disminuyó la diversidad tanto de las Poaceae C3 como de las C4.
Análisis de correspondencias
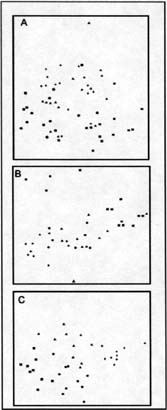
Al cabo del primer año de iniciados los tratamientos el DCA mostró claramente
los efectos del fuego, del pastoreo y de su combinación. Las dos comunidades
se ordenaron claramente a lo largo de los ejes de variación. El eje 1 se asoció
al fuego y el eje 2 al pastoreo (Figura 6). La situación excluída al pastoreo
y no afectada por la quema se situó en el extremo derecho del diagrama. Los
efectos combinados del fuego y el pastoreo direccionaron la comunidad hacia
una composición diferente respecto al resto de los tratamientos y tal combinación
permaneció diferenciada durante todo el ensayo (Figura 6 B y C). La exclusión
al pastoreo diluyó los efectos iniciales del fuego a los 3 años, pero ese período
de tiempo no fue suficiente para eliminarlos totalmente. La situación de referencia
(pastoreo sin fuego), no tuvo una identificación clara en el diagrama (Figura
6 C).
 |
|
| Figura 6: |
Ordenamiento CDA de los tratamientos experimentales en la comunidad de Festuca hieronimy al primer (A), segundo (B) y tercer (C) año. Tratamientos: Con pastoreo sin fuego (triángulos), sin pastoreo sin fuego (cruces), con pastoreo con fuego (cuadrados) y sin pastoreo con fuego (circulos). |
| Figure 6: |
CDA classification of the experi- mental treatments in the Festuca
hieronimy community during the first (A), second (B) and third (C) years.
Treatments: With grazing without fire (triangles), without grazing or fire
(crosses), with grazing and fire (squares) and without grazing with fire
(circles). |
Comunidad de Stipa juncoides Sorghastrum pellitum
Esta comunidad ocupa laderas de poca inclinación, repartiendose la cobertura
entre varias especies, tales como: Stipa juncoides, Sorghastrum pellitum,
Schizachyrium spicatum y Stipa filiculmis. La cobertura total no
sobrepasa el 60% y la altura promedio alcanza a 30 cm. Este pastizal no es invadido
por malezas alóctonas.
Diversidad y cobertura
La remoción del pastoreo incrementó la cobertura de las Poaceae C3
y C4 y cuando ella estuvo combinada con la ausencia de fuego, favoreció
sólo a las Poaceae C4 (Figura 7). El pastoreo no afectó la competencia
entre las Poaceae C3 y C4, y el fuego, contrariamente
a lo esperado, ejerció un efecto estimulante sobre el segundo grupo (Figura
7). La ausencia de fuego y remoción del pastoreo inicialmente incrementó la
cobertura de ambos grupos, pero al 3er año, la cobertura de las Poaceae
C4 se duplicó reduciendo la cobertura de las Poaceae C3
a la mitad de su valor inicial (Figura 7).
 |
|
| Figura 7: |
Cobertura de Poaceae C3 (barras negras) y C4 (barras blancas) en la comunidad de Stipa juncoides-Sorghastrum pellitum al primer (A) y tercer (B) año de ocurrido el fuego. |
| Figure 7: |
Cover of C3 Poaceae (black bars) and C4 Poaceae (white bars) in the
Stipa juncoides- Sorghastrum pellitum community during the first
(A) and the third (B) years after the fire. |
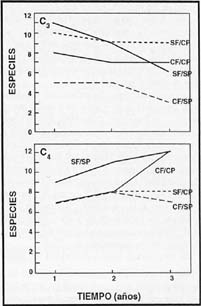
La presencia de fuego estuvo asociada a un incremento en la diversidad de la
comunidad, especialmente de las Poaceae C4, mientras que la ausencia
de pastoreo incrementó la diversidad de las Poaceae C3 (Figura 8)
. La condición sin fuego-sin pastoreo, fue la única donde la diversidad se mantuvo
con valores similares para ambos grupos (Figura 8). A excepción del tratamiento,
pastoreo sin fuego y desde el primer año de iniciados los tratamientos, la diversidad
de las Poaceae C3 fue superior a la del grupo C4. Al final
del ensayo, la diversidad de las Poaceae C4 fue superior en todos
los tratamientos, registrándose valores similares pero siempre mayores en el
tratamiento sin fuego + sin pastoreo y la mayor diferencia fue registrada para
el tratamiento pastoreo + fuego ( Figura 8).
 |
|
| Figura 8. |
Variación de la riqueza específica de especies de Poaceae C3 (arriba) y de Poaceae C4 (abajo) en la comunidad de Stipa juncoides-Sorghastrum pellitum en los tres primeros años. |
| Fihure 8: |
Variation of the species richness of C3 Poaceae (above) and C4 Poaceae
(below) in the Stipa juncoides-Sorghastrum pellitum community during
the first three years. |
Análisis de correspondencias
La localización de los tratamientos en el diagrama de ordenación tuvo un diseño
similar al del pastizal alto ( Figuras 6 y 9). Los efectos combinados del fuego
y el pastoreo direccionaron a la comunidad hacia otra composición florística
diferente al resto de los tratamientos, efecto que se mantuvo al cabo del tiempo
que duró la experiencia (Figura 9).
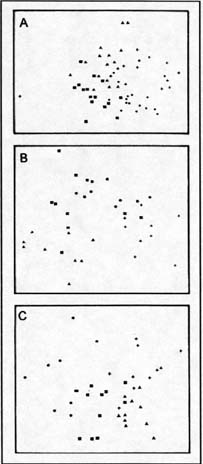 |
|
| Figura 9: |
Ordenamiento CDA de los tratamientos experimentales en la comunidad de Stipa juncoides-Sorghastrum pellitum al primer (A), segundo (B) y tercer (C) año. Tratamientos: Con pastoreo sin fuego (trángulos), sin pastoreo sin fuego (cruces), con pastoreo con fuego (cuadrados) y sin pastoreo con fuego (circulos). |
| Figure 9: |
CDA classification of the experimental treatments in the Stipa juncoides-Sorghastrum
pellitum community during the first (A), second (B) and third (C) years.
Treatments: With grazing without fire (triangles), without grazing or fire
(crosses), with grazing and fire (squares) and without grazing with fire
(circles). |
La ausencia de fuego diluyó los efectos del pastoreo y recíprocamente su presencia
y la remoción del pastoreo direccionó a la comunidad hacia otra composición
florística diferente a la del resto de los tratamientos (Figura 9 B y C), especialmente
a lo referido al sitio quemado y pastoreado.
La liberación del espacio y la presencia de suelo desnudo en el pastizal como
consecuencia del fuego estimuló la germinación y rebrote de las Poaceae C4,
incrementando la diversidad inicial de las mismas, efecto que se mantuvo a lo
largo de los 3 años de la experiencia. El fuego incrementó la cobertura y diversidad
de las Poaceae C4, resultados que concuerdan con lo hallado por Collin
y Wallace (1990) y su combinación con el pastoreo ejerció los efectos más
notables (Figuras 4 y 7).
Los resultados obtenidos indican que el cambio en la estructura de la vegetación
causado por el fuego no es transitorio (Noy Meir, 1995); y
por el contrario, los efectos interactivos del fuego y pastoreo pueden ser muy
deletéros, especialmente en la comunidad dominada por Poaceae C3
(Figuras 6 C y 9 C). Ello puede deberse a que el pastoreo postfuego selectivo
de las Poaceae dominantes C3, mejora la posición competitiva de las
especies C4, por encima de lo realizado por el fuego (Lubchencko,
1978; Ode et al., 1980) al cambiar la estructura del canopeo,
favoreciendo la entrada de luz.
En el pastizal bajo (comunidad de Stipa juncoides-Sorghastrum pellitum),
codominado por Poaceae C3 y C4, contrariamente a lo esperado,
los efectos combinados del fuego y pastoreo no ejercieron efectos significativos
sobre la cobertura respecto a la situación de referencia. Es posible que esto
se relacione a la propia identidad de estos sitios: tienen generalmente un canopeo
denso y bajo, son fuertemente pastoreados y el material combustible es muy escaso
(Mc Naugthon, 1984; Hobbs y Mooney, 1991)
La exclusión al pastoreo ejerció efectos contrastantes según su asociación con
fuego. Combinado con fuego niveló la competencia de las Poaceae C3
y C4, pero su ausencia favoreció a las Poaceae C4 (Figura
7). En esta comunidad, el pastoreo juega un rol importante ya que el mismo es
selectivo y el pasto vaca (Sorghastrum pellitum), es recurrentemente
pastoreado, ello se ve en el aumento llamativo de la cobertura de las Poaceae
C4 en la situación de no pastoreo. En el pastizal alto (comunidad
de Festuca hieronymi), los efectos del pastoreo quedaron atenuados luego
de 3 estaciones de crecimiento; la ausencia de fuego favoreció el crecimiento
de las Poaceae C3 que terminaron dominando el pastizal (Figura 4).
El pastoreo contínuo y liviano, que es la situación de manejo más practicada,
en general favorece a las Poaceae C3 de alto porte, que cuando adultas
son de baja palatabilidad (Figura 4).
Esta situación de manejo potencia la dominancia de las C3, cerrando
rápidamente el canopeo y evitando la llegada de luz al suelo, disminuyendo así
la posibilidad de germinación de plántulas. También favorece la acumulación
de material muerto que no es comido por el ganado, situación que actúa como
disparador de la decisión del productor, quien usualmente quema el pastizal
para obtener un rebrote de mejor calidad (Menghi et al., 1993).
El fuego ejerció un efecto estimulante sobre el crecimiento de las Poaceae C4
al modificar las condiciones microclimáticas del pastizal ( Ode
et al., 1980). La exclusión del pastoreo postfuego, posibilitó la expresión
de las Poaceae C4, pero fueron las Poaceae C3 las que
recuperaron más rápidamente la dominancia. La quema del pastizal con pastoreo
continuo y liviano, al año de ocurrido el evento, estimuló el crecimiento de
las Poaceae C3, sin embargo, las Poaceae C4, comenzaron
a crecer más vigorosamente al 2do y 3er año post-fuego,
beneficiadas por el pastoreo selectivo de las Poaceae C3 (Figura
4).
En el pastizal alto, el fuego y el pastoreo como eventos aislados benefician
a las Poaceae C4, pero cuando interactúan, las relaciones de competencia
se mantienen e impiden que las Poaceae C3 reestablezcan su dominancia
(Figura 4). Con ello se modifica la estructura: desaparece un pastizal alto
( más de 80 cm), de baja diversidad, típicamente invernal e induce hacia uno
más bajo (menor de 60 cm), más diverso y con modificación de la oferta forrajera
a través del año. Ello puede deberse a que la forma de crecimiento de los pastizales
altos, con una inversión relativa mayor en producción aérea a subterránea (Sims
y Singh, 1978) son adaptaciones que mejoran la competitividad por luz de
estos pastizales, pero a su vez los torna más vulnerables al pastoreo por los
grandes herbívoros especialmente luego del fuego, suceso que aumenta la diversidad
pero altera la oferta forrajera (Milchunas et al., 1988).
Los resultados obtenidos concuerdan con las afirmaciones de Howe
(1994), quien sostiene que en los pastizales altos la quema durante la estación
de reposo y la exclusión al pastoreo podrían promover artificialmente la dominancia
de las Poaceae C4. En el pastizal alto el fuego promueve el crecimiento
y la diversidad de las Poaceae C4, y el pastoreo mantiene las relaciones
de competencia, mientras que en el pastizal bajo quien promueve el crecimiento
de la Poaceae C4 es la remoción del pastoreo. El conocimiento de
la respuesta de estos grupos funcionales es de suma importancia para prescribir
normas de manejo en estos pastizales muy heterogéneos. Además, dada la disponibilidad
de espacio en ambos pastizales, la posibilidad de autoresiembra debe ser alta,
por lo que a futuro será necesario hacer estudios del banco de semillas, tanto
activo como pasivo, para determinar su grado de influencia en la composición
botánica (Harper, 1977).
Es necesario investigar sobre otros métodos de pastoreo, ya que la interacción
fuego-pastoreo altera la estructura del pastizal y los rezagos parecen ser muy
importantes en la comunidad de pastizal bajo.
Se impone, entonces la necesidad de continuar el estudio con otras comunidades
representativas del pastizal con el objetivo de ampliar las bases del conocimiento
que permitan sustentar un manejo que compatibilice las necesidades de los productores
del lugar (interés privado) con las necesidades de preservación de biodiversidad
(interés público).
De los resultados descritos se pueden extraer las siguientes conclusiones:
- Los grupos funcionales, de Poaceae C3 y C4 fueron indicadores
adecuados para evaluar las respuestas del pastizal a los disturbios y permitió
entender que la combinación fuego y pastoreo debe utilizarse, considerando los
efectos particulares que ejercen sobre cada comunidad, lo cual demanda un monitoreo
de los cambios operados por los grupos funcionales si se quiere realizar un
manejo racional del ecosistema pastizal.
- El uso del fuego como única herramienta de manejo, para mejorar el componente
herbáceo C4 de la comunidad de los pastizales altos no parece factible.
En el pastizal alto el uso selectivo de las Poaceae C3 por los herbívoros
luego de la quema, mejora la posición competitiva de las Poaceae C4,
pero no sucede lo mismo en el pastizal bajo.
- El sistema de pastoreo continuo y extensivo, practicado desde hace muchos
años por los ganaderos de la región, no puede ser aplicado de la misma forma
a las dos comunidades, especialmente cuando se lo combina con el fuego. En el
pastizal alto los efectos del pastoreo postfuego ejercen efectos que modifican
los atributos del pastizal y se podría esperar grandes modificaciones de seguir
las normas de manejo actual, situación que no ocurre con el pastizal bajo.
-Es importante considerar el comportamiento diferencial de las 15 comunidades
que integran el sistema de pastizales naturales serranos. Aquí se expresan las
conclusiones del estudio de solamente dos de ellas ; ambas difieren en sus respuestas
al fuego y al pastoreo.
ANDERSON, R.C. 1982. An evolutionary model summarizing the roles of fire, climate and grazing animals in the origin and maintenance of grasslands: an end paper. In: J. R. Estes, R. J. Tyrl and J. N. Brunken (eds), Grasses and Grasslands: Systematic and Ecology, pp 297-308. Univ. Oklahoma Press., Norman. OK.
BELSKY, A. J. 1992. Effects of grazing, competition, disturbance and fire on species composition and diversity in grassland communities. J. Veg. Sci. 3: 187-200.
BIANCO, C. , KRAUS, T., ANDERSON, D. y CANTERO, J. 1987. Formaciones vegetales del suroeste de la provincia de Córdoba. Rev. UNRC. 7 (1): 5-66.
CABIDO, M., ATECA, N., ASTEGIANO, M. E. y ANTON, A. M. 1997. Distribution of C3 and C4 grasses along an altitudinal gradient in Central Argentina. J. of Biogeograp. 24: 197-204.
CANTERO, J. J. y BIANCO, C. A. 1986. Las plantas vasculares del suroeste de la provincia de Córdoba (Argentina), Parte III. Catálogo preliminar de las especies. Rev. UNRC 6 (1): 5-52.
CAVAGNARO, J. B. 1988. Distribution of C3 and C4 grasses at different altitudes in a temperate arid region of Argentina. Oecología 76: 273-277.
COLLINS, S. L. 1987 Interactions of disturbances in tallgrass prairie: A field experiment. Ecology 68: 1243-1250.
COLLINS, S. L. y WALLACE, L. C. 1990. (eds), Fire in North American Tallgrass Prairies. University of Oklahoma Press, Norman, Oklahoma. 308 pp.
DEREGIBUS, V. A., SANCHEZ, R. A., CASAL, J.J. y TRLICA, M. 1985. Tillering responses to enrichment of red light beneath the canopy in a humid natural grassland. J. of Appl. Ecol. 22:199.206.
DIAZ, S., ACOSTA, A. y CABIDO, M. 1994. Grazing and phenology of flowering and fruiting in a montane grassland in Argentina: a niche approach. Oikos 70: 287-295.
DÍAZ, S. y CABIDO, M. 1997. Plant functional types and ecosystems function in relation to global change. J. Veg. Sci. 8: 464-474.
EPSTEIN, H. E., LAUENROTH, W. K., BURKE, I. C. y COFFIN, D. P. 1997. Productivity patterns of C4 and C3 functional types in the U.S. great plains. Ecology, 78(3): 722-731.
GASTON, K. J. 1996. Species richness: measure and measurement. In: Gaston, K. J. (ed). Biodiversity: a biology of numbers and difference, pp. 77-113. Blackwell Science, Oxford.
GONZALEZ BONORINO, F. 1950. Algunos problemas geológicos de las Sierras pampeanas. Raga 5: 81-110.
GORDILLO, C. y LENCINAS, A. 1979. Geología Regional Argentina. Acad. Nac. Cienc. Córdoba. 463 pp.
GREIG-SMITH, P. 1964. Quantitative plant ecology. Butterworths, Londres. 2da. edición, 256 págs.
HARPER, J.L. 1977. Population biology of plants. Academic Press, Londo, New York, San Francisco. 892 pp.
HOBBS, R. J. y MOONEY, H. A. 1991. Effects of rainfall variability and gopher disturbance on serpentine annual grassland dynamics. Ecology 72: 59-68.
HOWE, H. F. 1994. Managing species diversity in tallgrass prairie: Assumptions and implications. Conser. Biolol., 8 (3): 691-704.
INSTITUTO NACIONAL DE TECNOLOGÍA AGROPECUARIA (INTA) y Ministerio de Agricultura, Ganadería y Recursos Naturales Renovables (MAGYRN). 1994. Carta de suelos de la República Argentina. Hoja 3366-12 Río de los Sauces y Hoja 3366-18 Alpa Corral.
KETLING, R. W. 1954. Effects of moderate grazing on the composition and plant production of a native tall-grass prairie en central Oklahoma. Ecology 35: 200-207.
KURTZ, F. 1904. Flora. Vol I: 270-343. In M. Río y Achával (eds.). Geografía de la Provincia de Córdoba., Buenos Aires.
LUBCHENCKO, J. 1978. Plant species diversity in a marine intertidal community: importance of herbivore food preference and algal competitive abilities. Am. Nat. 112: 23-29.
LUTI, R., BELTRAN, M., GALERA, F., FERREIRA, N., BERZAL, M., NORES, M., HERRERA, A. y BARRERA, J. 1974. Vegetación. En: Vasquez, J.; Miatello, R. y Roque, E. (eds.). Geografía física de la provincia de Córdoba. Fund. Bco. Prov. Córdoba. 463 pp.
MARTINO, R., KRAEMER, P., ESCAYOLA, M., GIAMBASTIANI, M. y ARSONIO, M. 1995. Transecta de las Sierras Pampeanas de Córdoba a los 32° S. RAGA 50: (1-4). 40-47.
Mc CUNE, B., y MEFFORD, M. J. 1999. PC-ORD. Multivariate Analysis Ecological Data, Version 4, MjM Software Design, Gleneden Beach, Oregon, USA. 237 pp.
MC NAUGHTON, S.J. 1984. Grazing lawns: Animals in herds, plant form, and coevolution. Am. Nat. 124: 863-885.
MENGHI, M., M. CABIDO, A. ACOSTA, B. PECO y F. D. PINEDA 1993. Changes in pasture communities subjet to burning in the Córdoba mountains, Argentina. Coenoses 8 (1):1-10.
MILCHUNAS, D. G. , O. E. SALA y W. K. LAUENROTH. 1988. A generalizad model of the effects of grazing by large herbivores on grassland community structure. Am .Nat. 132: 87-106.
NOBLE, I. R. y GITAY, H. 1996. A functional classification for predicting the dynamics of landscapes. J. Veg. Sci. 7:329-336.
NOY-MEIR, I. 1995. Interactive effects of fire and grazing on structure and diversity of Mediterranean grasslands. J. Veg. Sci. 6: 701-710.
ODE, D. J., L. L. TIEZEN, y J. C. LERMAN. 1980. The seasonal contribution of C3 and C4 plant species to primary production in a Mixed Prairie. Ecology 61: 1300-1311.
PARODI, L. E. 1964. Regiones fitogeográficas argentinas. Enciclopedia de Agricultura y jardinería. Tomo II. Ed. Acme. Buenos Aires. 1161 pp.
PARUELO, J. M., JOBBÁGY, E. G., SALA. O. E., LAUENROTH, W. K. y BURKE, I. C. 1998. Functional and structural convergence of temperate grassland and shrubland ecosystems. Ecol. Appl. 8 (1): 194-206.
PEARCY, R. W. y EHLERINGER, J. 1984. Comparative ecophysiology of C3 and C4 plants. Plant, Cell and Environment 7:1-13.
PENFOUND, W. T. 1964. The relation of grazing to plant succesion in the tall grass prairie. J. Range Management 17: 256-260.
POLLEY, W. N. 1997 Implications of rising atmospheric carbon dioxide concentration for rangelands. J. Range Management 50(6): 561-577.
RAGONESE, A. 1967. Vegetación y ganadería de la República Argentina. Colección Científica INTA : 5. Argentina. 218 pp.
SALA, O. E. 1988. The effect of herbivory on vegetation structure In. M. J. A. Werger, P. J. M. van der Aart, H. J. During y J. T. A. Verhoeven. (eds). Plant Form and Vegetation Structure, pp 317-330. SPB Academic Publishing, The Hague, Netherlands.
SÁNCHEZ, E. y ARRIAGA, M. O. 1990. El síndrome kranz en Poaceae de la Flora Argentina. Parodiana, 6(1): 73-102.
SIMS, P. L. y J. S. SINGH. 1978 The structure and function of ten western North American grasslands. III. Net primary production, turnover and efficiences of energy capture and water use. J. Ecol. 66: 573-597.
STEUTER, A. A. 1987 C3/C4 production shift on seasonal burns-northern mixed prairie. J. Range Management 40 (1): 27-31.
TILMAN D. y DOWNING J. A. 1994. Biodiversity and stability in grasslands. Nature 367 : 363-365.
WALKER, B. H. 1992 Biodiversity and ecological redundancy. Conserv. Biol. 6: 18-23.
ZULOAGA, F. O., NICORA, E. G., RÚGOLO DE AGRASAR, Z. E., MORRONE, O., PENSIERO, J. y CIALDELLA, A. M. 1994. Catálogo de la familia Poaceae en la República Argentina. Monogr. Syst. Bot. Missouri Bot. Gard. 47:1-178.