CONTROL POTENCIAL DEL COLOR CAUSADO POR EL DESARROLLO DE
MAGULLADURA EN PAPAS RANGER RUSSET MEDIANTE ALMACENAMIENTO EN ATMOSFERA CONTROLADA
Loyola, N.ï,
Thornton, R.E. *, Hiller, L.K. * Y Powers, J‡.
ï Universidad Católica del Maule,
Escuela de Agronomía. Carmen 684 Curico, VII región Chile. *Horticulture
and Landscape
Architecture Dept., ‡Food Science and Human
Nutrition Dept., Washington State University.
Recepción originales: 13 de junio del
2002
ABSTRACT
The Potential of Controlled Atmosphere Storage to Control
Tuber Bruise Development in Ranger Russet Potatoes.
The potential for controlled atmosphere (CA) storage to maintain
quality of Ranger Russet potato (Solanum tuberosum L.) tubers was investigated.
Before storage condition tests were initiated (time zero), potatoes were divided
into two groups, bruised and non-bruised tubers. Tubers were stored (6.6 ¥C,
95% RH) in regular atmosphere (RA), 5% CO2 12% O2
or 5% CO2 6% O2. The assessment
of tuber response to storage conditions included the evaluation of bruised and
non-bruised tuber flesh color after two months of storage. Color was measured
immediately after peeling, 10 and 60 minutes later, and after 24 h. The flesh
color of bruised tubers, compared to non-bruised tubers at time zero, showed
significant changes in lightness (L*), but not in green coloration (a*) or yellowness
(b*). The bud and stem ends were equally susceptible to bruise effects as reflected
by the parameters L*, a* and b*. After two months storage, RA and 5% CO2
6% O2 were the best conditions to reduce the degree
of darkness in bruised tubers. The 5% CO2 6% O2
atmosphere reduced changes in a* values, but did not affect b* values.
Keys words: Ranger Russet, controlled
atmosphere, bruise, color.
RESUMEN
Se investigó la capacidad de dos atmósferas controladas
(AC) para mantener la calidad de papas Ranger Russet (Solanum tuberosum L.).
Antes de iniciar las condiciones de almacenamiento (tiempo cero), las papas
fueron divididas en dos grupos, tubérculos con y sin magulladuras. Los
tubérculos fueron almacenados (6,6 ¥C,
95% HR) en atmósfera regular (AR), en 5%CO2
12%O2 o en 5%CO2 6%O2.
La respuesta de los tubérculos a las condiciones de almacenamiento, incluyó
la evaluación del color de la pulpa de los tubérculos, con y sin
magulladura, luego de dos meses de almacenamiento. El color fue medido inmediatamente
después del pelado, 10 y 60 minutos mas tarde, y después de 24
h. El color de la pulpa de tubérculos magullados, comparado con los no
magullados al tiempo cero, exhibió cambios significativos en luminosidad
(L*), pero no en el color verde (a*) ni en el amarillo (b*). La yema y tallo
apicales, fueron igualmente susceptibles al efecto de la magulladura, según
se reflejó en los parámetros L*, a* y b*. Después de dos
meses de almacenamiento, la AR y la con 5%CO2 6%O2
fueron las mejores condiciones para reducir el grado de oscurecimiento en tubérculos
magullados. La atmósfera de 5%CO2 6%O2
redujo los cambios en valores de a*, pero no afectaron los valores de b*.
INTRODUCCION
Las pérdidas de papas durante la cosecha y la manipulación
son aproximadamente de un 8% (USDA, 1990) con la magulladura
por el daño por impacto, como la mayor contribución a esta pérdida.
Papas con magulladura significan un costo adicional, debido a las labores extras
de selección para reemplazar el material dañado y sin uso industrial
(USDA, 1994). Se estima que en EE.UU., cada año, llegan
a la industria procesadora de papas con magulladuras que representan entre el
20 y el 30% del valor total y que un 40% del cultivo tiene al menos un 5% extra
de reducción en tamaño por magulladura (USDA, 1994).
En relación a las propiedades físicas de los
tubérculos, Wendell (1995) reportó que la muestra
de tejidos (sector de yemas y tallos), puede permitir una acuciosa evaluación
de las propiedades físicas, las cuales pueden ser usadas para predecir
la tendencia en la sensibilidad al impacto, pero no puede medir directamente
la sensibilidad al impacto de todo el tubérculo, ello debido a las diferencias
geométricas y anatómicas de los tubérculos, tales como;
desarrollo de tejido vascular y grosor de la corteza.
Mucho de los daños ocurren por impactos durante la
cosecha, manipuleo y comercialización, como consecuencia de que los tubérculos
se ponen en contacto con partes del equipo y también con piedras (Kadam
et al., 1991). Según Mohsenin (1986), la magulladura
se define como daño en el tejido de las plantas, debido a fuerzas externas
y que resultan en un cambio en la textura del tejido y en el color. Las alteraciones
físicas o daños mecánicos, pueden ser divididos en dos
grupos: Magulladuras internas o «black spot» y daños externos,
lo que incluye daño en la piel en la forma de machucadura, ablandamiento
y roturas (Gray y Hughes, 1978; Kadam et al.,
1991). La magulladura interna con desmenuzamiento, fue descrita por Noble
(1985), reconociendo un círculo o forma de estrella, como resultado
por impacto a gran velocidad. Este autor también describió la
machucadura interna, como un daño causado por impacto a baja velocidad.
Todo tipo de daño mecánico y físico está influido
por factores tales como cultivares, grado de madurez, estado de crecimiento
y condiciones de almacenamiento (Gray y Hughes, 1978).
El color oscuro asociado con la magulladura negra o «black
spot bruise», está relacionado con el desarrollo de melanina en
el tejido (Zhang et al., 1989). Los impactos del acopio, causan
rotura de las membranas celulares y permiten que la enzima polifenol oxidasa
(PPO) y el sustrato tirosina se pongan en contacto, resultando la oxidación
de este ultimo para formar melanina, en muchos casos sin destrucción
de las paredes celulares (Zhang et al., 1989). La hidroxilación
de la tirosina, resulta en la producción de dihidroxifenil-alanina (DOPA)
y luego en una rápida oxidación de DOPA a DOPA quinona. La DOPA
quinona, combina la oxidación a compuestos rojos cromofóricos
y últimamente a melanina. El pardeamiento resultante, gris, o tonos negros
de los tejidos magullados, está relacionado con la cantidad de la melanina
presente (Zhang et al., 1989). Sawyer y Collin
(1960) establecieron que el potasio, tiene una influencia en las magulladuras
y manchas negras y que probablemente afecta indirectamente la gravedad especifica
de los tubérculos. Hay una influencia del cultivar, por ejemplo: Ranger
Russet, es mas susceptible al magullamiento negro que Russet Burbank, Kennebec
y Norchip. El ultimo, es considerado como resistente al magullamiento negro
(Blight y Hamilton, 1974, Pavek et al., 1992).
Estudios realizados con tubérculos inmaduros y que
presentaron magulladura negra o «blackspot bruise», sufrieron un
control moderado en atmósferas de (3 a 5)% CO2
y aire (Timm et al., 1976). La medición del color de
la magulladura, es una combinación de valores de L*, a*, y b* (Hunter
Associates Laboratory, 1991). La mayoría de la investigación
sobre magulladura en almacenamiento, tiene en consideración el desarrollo
de magulladuras en relación al cambio de color, actividad enzimática,
composición atmosférica normal del aire, influencia del etileno,
y temperatura de almacenamiento. El presente estudio, relaciona almacenamiento;
temperatura, tipos de atmósferas y tiempo para concluir que la AR y 5%CO2
6%O2, fueron las mejores condiciones para el control
del fenómeno de magulladura en el almacenamiento de papas cv. Ranger
Russet.
La hipótesis de la presente investigación, es
que la cantidad de polifenol oxidasa, responsable del oscurecimiento en tubérculos
magullados, podría ser bloqueada con incremento en los niveles de CO2
y disminución en los niveles de O2, lo que
podría atenuar la manifestación de magulladuras durante el almacenamiento.
La presente investigación, tuvo como objetivo principal,
evaluar el efecto de dos condiciones de almacenamiento en atmósferas
controladas: (5%CO2 12%O2
y 5%CO2 6%O2) y atmósfera
regular (0,03%CO2 21%O2),
sobre el pardeamiento enzimático de tubérculos de papas magulladas
y almacenadas por dos meses.
MATERIALES Y METODOS
Se curaron tubérculos de papas (Solanum tuberosum) cv.
Ranger Russet en bolsas plásticas con 14 tubérculos entre 227g
y 340g por malla a 8,8¥C
y 95% HR por 30 días , antes de almacenarlos en atmósferas controladas.
Después del curado, las mallas con papas fueron divididas aleatoriamente
en tres lotes y ubicadas en las siguientes tres condiciones de almacenamiento:
0,03%CO2 y 21%O2 (AR), 5%CO2
12%O2 y 5%CO2 6%O2.
Todos los tubérculos, fueron mantenidos en esas condiciones a 6,6¥
C y 95% HR por 2 meses. La condición de AR, fue obtenida utilizando la
composición del aire al interior de la principal cámara de mantención.
La condición de AC, a través de un sistema continuo, fue aplicada
a los tubérculos mantenidos en contenedores de plexiglas, ubicadas al
interior de la cámara de mantención. La tasa de inyección
de gas al interior de las cámaras de plexiglas de 180 L fue: 79,1 mL
N2/min, 110,9 mL aire/min. y 10 mL CO2/min
para la condición de almacenamiento de 5%CO2
12%O2 y 138,8 mL N2/min,
51,2 mL aire/min, 10 mL CO2/min para la condición
de 5%CO2 6%O2. La tasa de
gas al interior de cada atmósfera controlada fue verificada cada 8 horas,
al igual que la emisión de CO2 fuera de las
cámaras de plexiglas. La humedad fue provista por agua en los bordes
superiores de las cámaras de plexiglas y que sirvió además
de sellador. El estudio fue además, dividido en dos experimentos.
1. Evaluación del Color de Papas Magulladas
en el Tiempo Cero de Almacenamiento.
Los tubérculos curados, fueron divididos en dos subgrupos: uno destinado
a ser magullado y el otro para servir como control sin magullar. Cada tubérculo
fue marcado con una marca de lápiz permanente, en los sectores de yema
y tallo en ambos lados. La mitad de los tubérculos, fueron sostenidos
manualmente y magullados mediante un instrumento de impacto (modificado por
Johnson, 1972), constituido por una esfera de 113g ubicada
en un extremo de un tubo estacionario de 58,4 cm. que impacta las papas en las
zonas marcadas.
Después de pelar manualmente los tubérculos,
se midió el color de la pulpa en las zonas marcadas de cada subgrupo
en el tiempo cero, inmediatamente después de peladas, 10 y 60 minutos
mas tarde y después de 24 h., utilizando directamente una cámara
Minolta chroma Meter CR-200 (Minolta Co., Japan), sobre las áreas marcadas
de las papas. La lectura, según los datos recolectados desde 8 tubérculos
por muestra, fueron ingresados en un procesador de datos DP-200 (Hunter
Associates Laboratory, 1991).
2. Evaluación del Color en Papas Magulladas
Después de Dos Meses de Almacenamiento.
Papas de ambos sub-grupos, magulladas y sin magullar, fueron almacenadas durante
dos meses en las tres condiciones de almacenamiento (AR, 5%CO2
12%O2 y 5%CO2 6%O2).
Transcurridos dos meses de almacenamiento, el color de la pulpa de papas con
y sin magulladuras se midió inmediatamente después de pelado,
10 y 60 minutos y 24 horas mas tarde. El color se determinó, según
se describió para el grupo de papas al tiempo cero de almacenamiento.
Se utilizó, para ambos estudios, un diseño de
bloques completos al azar con arreglo factorial y tres repeticiones (Condiciones
Almacenamiento x Magulladura x Parte del Tubérculo x Tiempo de Almacenamiento).
Los datos fueron analizados por ANDEVA con una significancia de p < 0,05
según SAS (SAS Institute Inc., 1991) y una separación
de medias según Duncan (Ott, 1992).
RESULTADOS Y DISCUSION
Evaluación del Color de Papas Magulladas en
el Tiempo Cero de Almacenamiento.
La magulladura tuvo efectos significativos en los valores L* (Cuadro
1). Los tubérculos sin magulladuras, tanto al tiempo cero, como en
todo momento exhibieron más bajos valores de L* (color oscuro de la pulpa)
que aquellos tubérculos magullados. Esto se debe, probablemente al hecho
de que los últimos, necesitan mas de 24 h para mostrar cualquier cambio
importante (Hudson, 1975). Los valores de L* en tubérculos
magullados (70,98) y no magullados (68,02) fueron estadísticamente diferentes,
pero estaban en el mismo rango de color de acuerdo a Hunter Associates
Laboratory (1991).
Cuadro 1. Influencia de la magulladura,
parte del tubérculo, y tiempo después de pelado de los tubérculos
sobre los valores de L*, a*, y b* en papas cv. Ranger Russet al inicio
de la investigación.
Table 1. Effect of bruising, location in tuber, and time
after peeling, on L*, a* and b* values on potato cultivar Ranger Russet
at the start of the experiment. |
| |
Fuente |
Luminosidad
L* |
a* |
b* |
|
| Magullado |
70,98 A^ |
3,43 A |
20,23 A |
| No-magullado |
68,02 B |
3,63 A |
20,76 A |
| Yema |
69,43 A |
3,57 A |
20,50 A |
| Tallo |
69,57 A |
3,49 A |
20,49 A |
|
| Tiempo después de pelado(valores promedios en tubérculos
magullados y no magullados) |
0 |
74,57 A |
4,21 A |
22,69 A |
10 min |
74,51 A |
4,11 A |
22,29 A |
60 min |
72,53 B |
3,71 B |
21,77 A |
24 hr |
56,39 C |
2,10 C |
15,24 B |
|
|
| ^ Promedios con la misma letra en una columna, no son
significativamente diferentes, según la prueba de Duncan al 5% de
probabilidad. |
Los tubérculos magullados, fueron afectados por daños
físicos en las células, las cuales perdieron turgor, lo cual pudo
provocar que la luz se reflejara al momento de recolectar los datos por la cámara
fotográfica. Aquel fenómeno, puede ser explicado por los tubérculos
magullados que exhibieron un alto valor de L* en relación con los no
magullados, previamente a la aplicación de las tres condiciones de almacenamiento.
El efecto de magulladura, se explica por el cambio en el valor de L*, desde
luminoso o brillante a opaco según el daño evidente, más
que cambios en a* y b* (Cuadro 1).
Los resultados se obtuvieron utilizando tubérculos
completos, lo que es mas aplicable a la situación industrial, pero limita
la posibilidad de medir las propiedades físicas fundamentales tales como:
fallas frente al stress y esfuerzos provocados por las dificultades debidas
a la amplia variación de formas entre tubérculos del mismo tamaño
y cultivar (Wendell, 1995). Los valores de L*, a* y b* disminuyeron
con el tiempo después del pelado. Los valores tanto de L* como de a*,
fueron significativamente diferentes a los 60 minutos y 24 horas después
de pelado, comparados con 0 y 10 minutos luego del pelado (Cuadro
1). Los valores de b*, fueron diferentes solo a las 24 h. El efecto de la
magulladura y tiempo después de pelado, fue la principal variable que
afectó el cambio en color percibido por los valores de L*.
Los tubérculos magullados y no magullados, evaluados
antes de someterlos a las diferentes condiciones de almacenamiento, exhibieron
el mismo valor de L* desde 0 a 60 minutos después de pelados y luego
de 24 h de pelados se tornaron oscuros (Cuadro 1). Estos resultados,
demostraron que el desarrollo del color ocurrió localmente y después
de 24 h de magullados los tubérculos. El almacenamiento en AR, considerando
el oxígeno disponible, podría evitar que la enzima polifenoloxidasa
reaccione con el sustrato tirosina dando a los tejidos una apariencia oscura,
la cual se podría intensificar con el tiempo después de ocurrida
la magulladura y el pelado. Tubérculos magullados y no magullados, mostraron
el mismo color oscuro y tendencia en los valores de L* lo que puede ser responsable
de enmascarar el color verde, hasta ser observado como gris tanto en yemas como
en tallos (Cuadro 1).
Los tubérculos magullados y no magullados, exhibieron
la misma tendencia y rango de los valores de a* (Cuadro 1).
El valor de a*, disminuyó rápidamente durante el periodo entre
10 a 60 minutos después que los tubérculos se pelaron, tanto los
magullados, como sin magullar. Después de 60 minutos, luego de pelados
los tubérculos, se observó la aparición de un tono gris
relacionado con la oxidación enzimática del sustrato tirosina,
susceptibilidad a la magulladura y saturación del substrato. El grado
de tono verde, (valor a*) tanto en yemas, como en tallos de los tubérculos,
fue afectado en forma similar por el tiempo después de magullado (Cuadro
1). Sesenta minutos después del impacto físico, el valor de
a* para las porciones de yema y tallo, estuvo en el rango de tono gris.
La tolerancia de los tejidos a la magulladura, podría
cambiar según la ubicación en el tubérculo, composición
físico - química de las yemas y tallos (Mathew,
1992). Sin embargo, no se observó ninguna diferencia en las respuestas
de yemas y tallos en el presente experimento.
Loyola (1997) evaluó en papas que
la sección de los tallos, tenía más azucares reductores
que la sección de las yemas y que la cantidad decrecía empleando
una AR y la condición 5%CO2 6%O2
de almacenamiento. Es posible que en ambos sectores del tubérculo, la
conformación física de la pulpa, la cual incluye gránulos
de almidón y paredes celulares, no sea suficientemente fuerte para resistir
el impacto de la magulladura. La firmeza y el turgor celular, son factores físicos
esenciales relacionados con la magulladura, conocida como “black spot
bruising”. Las diferencias en la estructura de las lenticelas y estructura
epidermal en cada cultivar, contribuyen a la susceptibilidad de los tubérculos
(Sawyer and Collin, 1960). De acuerdo a observaciones, realizadas
en el presente estudio, el color oscuro en tubérculos magullados, podría
también ser dependiente de las variaciones en su forma , lo que podría
afectar la superficie impactada con el consecuente daño celular. La intensidad
del color oscuro (bajo valor L*), observado después de magullar las papas,
fue mas representativo que los valores de a* y b* medidos en las secciones de
yemas y tallos de los tubérculos. El rango del valor a*, existente entre
60 minutos y 24h, fue corroborado por las observaciones visuales realizadas.
Se encontraron diferencias significativas entre los valores L*, a*, y b* y el
tiempo después de pelado (Cuadro 1). El efecto visual
de la magulladura es un oscurecimiento de los tejidos afectados (bajo valor
L*), de esta forma, el rango de color desde café a gris, producto del
pardeamiento cubrió el color natural de la papa y ocultó los componentes
a* y b* de la escala de colores.
Tubérculos Almacenados por 2 Meses, Magullados
y Evaluado su Color a los 0, 10, 60 Minutos y 24 Horas más tarde.
Tubérculos magullados y no magullados, se almacenaron por un período
de dos meses, bajo las tres condiciones de almacenamiento, al cabo del cual
de midió el color, representado por L*, a*, y b, después de pelados,
10 y 60 minutos, y 24 horas mas tarde.
Después de 2 meses de almacenamiento, las variables
independientes, condición de almacenamiento, magulladura, parte del tubérculo
y tiempo después de pelado afectaron significativamente los valores L*,
a*, y b* (Cuadro 2). El rango de valores de L* y b* fue similar
a los valores reportados por los tubérculos evaluados primeramente, antes
del almacenamiento (Cuadro 1). El tiempo de almacenamiento,
pudo ser insuficiente para observar diferencias importantes, entre los tubérculos
magullados y los no magullados.
Cuadro 2. Influencia de las condiciones
de almacenamiento, magulladura y no magulladura, parte del tubérculo,
y tiempo después del pelado de los tubérculos, sobre los
valores de L*, a*, y b* en papas cv Ranger Russet.
Table 2. Effect of storage conditions , bruising , location
in tuber, and time after peeling , on L*, a* and b* values on potato cultivar
Ranger Russet. |
| |
Fuente
|
Luminosidad |
|
| |
L* |
a* |
b* |
|
Atmosfera Regular |
67,97 A^ |
- 2,08 B |
24,16A |
5 % CO2 12 %
O2 |
66,60 B |
- 1,33 A |
21,30B |
5 % CO2 6 % O2 |
67,26 AB |
- 1,37 A |
21,18B |
Magulladura |
65,36 B |
- 0,98 A |
22,65A |
No magulladura |
69,19 A |
- 2,21 B |
21,78B |
Yemas |
68,57 A |
- 2,00 B |
22,99A |
Tallos |
65,98 B |
- 1,18 A |
21,43B |
|
| Tiempo después de pelado(valores promedios de tubérculos
magullados y no magullados) |
0 |
72,36 A |
- 4,43 D |
24,10AB |
‘10 min |
71,35 B |
- 2,95 C |
24,52A |
60 min |
69,88 C |
- 1,73 B |
23,47B |
24 hr |
55.50 D |
2,74 A |
16,76C |
|
|
^ Promedios con la misma letra en la columna, no son significativamente
diferentes, según la prueba de
Duncan al 5% de probabilidad. |
Los tubérculos provenientes de las tres condiciones
de almacenamiento, evaluados desde inmediatamente después del pelado
hasta 24 h más tarde, exhibieron diferencias significativas en los valores
de L*, a* y b* y un progresivo cambio durante el tiempo después de pelado
(Cuadro 2). Sin embargo, la variación más sensible,
afectada por la reacción química de los tubérculos magullados
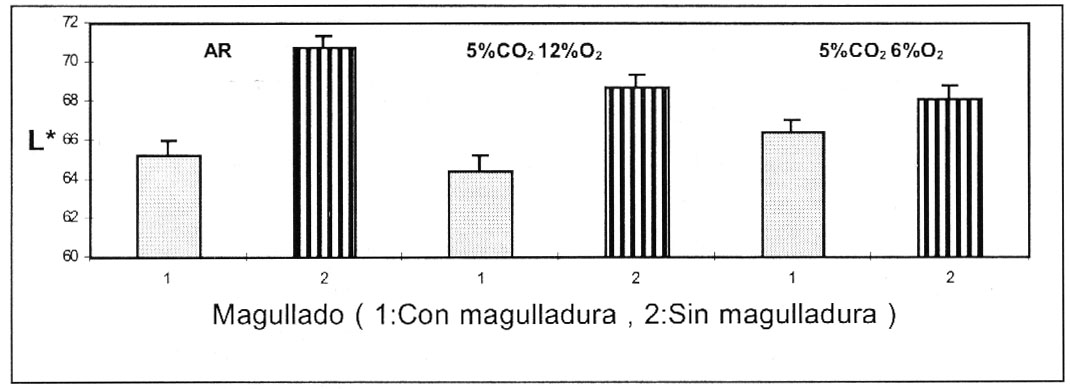
fue la luminosidad (valor L*). La figura 1, muestra el efecto
de la interacción entre las condiciones de almacenamiento y el magullado
en el oscurecimiento (valor L*) de los tubérculos magullados. Los tubérculos
almacenados en AR y en 5%CO2 6%O2
tuvieron un grado menor de oscurecimiento de su pulpa (alto valor de L*), que
aquellas pulpas de tubérculos almacenados en 5%CO2;
12%O2. Los tubérculos sin magullar, exhibieron
mayores valores de L*, almacenados en AR que aquellos almacenados en ambas condiciones
de AC.
|
| |
Figura 1. Luminosidad (valor L*)
de pulpa de papas cv Ranger Russet con y sin magulladuras, medidas 2 meses
después que se han introducido en las 3 atmósferas. Almacenamiento
a 6.6 ºC y 95 % HR
Figure 1. Luminosity (L* value) of flesh from potato
cultivar Ranger Russet with and without bruising, determined after 2 months
of storage under 3 conditions. Storage at 6.6 ºC and 95% RH. |
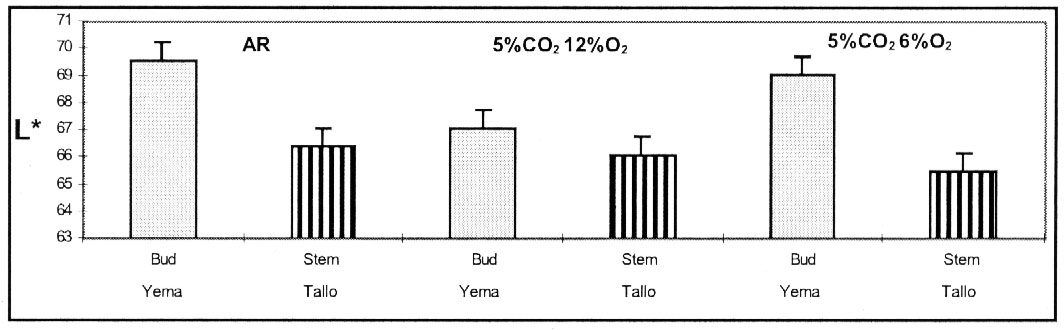
El tejido con magulladura, proveniente de sectores del tallo
de tubérculos fue igualmente oscuro, en las tres condiciones de almacenamiento
(Figura 2). El sector de las yemas, de tubérculos almacenados
en una atmósfera de 5% CO2 12%O2,
exhibió un color más oscuro, que el sector de las yemas de los
tubérculos provenientes de las otras dos condiciones de almacenamiento.
La cantidad de azúcares totales, azucares reductores y sacarosa acumulada
en los tubérculos almacenados en una atmósfera de 5%CO2
12%O2, puede haber afectado el balance de carbohidratos
en el sector de las yemas, resultando en una menor resistencia al impacto y
posterior magulladura con un bajo valor de L* (Loyola, 1997).
Dwelle y Stallknecht (1976), observaron un incremento en el
oscurecimiento de las partes magulladas, en tubérculos del cultivar Russet
Burbank, almacenados por un mes cuando la temperatura de almacenamiento fue
sobre los 10¥C.
Dichos autores, también reportaron que la temperatura óptima para
desarrollar el desorden, llamado “blackspot bruise”, varía
de 36¥C
a 40¥C
y observaron que a esas temperaturas las presiones de CO2
y N2, favorecen la formación de melanina, pero
a temperatura ambiente la presión del O2 no
tiene efecto significativo sobre el desarrollo de “blackspot bruise”.
La composición de los gases, en la atmósfera, fue más efectiva
para controlar este problema (“blackspot bruise”), si la temperatura
de almacenamiento fue menor a 10 ºC.
|
| |
Figura 2. Luminosidad (valor L*)
de pulpa de papas cv. Ranger Russet de porción de yemas y tallos,
medidos 2 meses después de iniciado el almacenamiento en 3 atmósferas.
Almacenados a 6.6 ºC y 95 % HR.
Figure 2. Luminosity (L* value) of bud and stem flesh
from potato cultivar Ranger Russet determined after 2 months of storage
under 3 conditions. Storage at 6.6 ºC and 95% RH. |
En el presente estudio, las dos condiciones de almacenamiento
de AC, parte del tubérculo, y el tiempo después de pelado, aportaron
más detalles acerca del comportamiento del cultivar Ranger Russet después
de magullados y el desarrollo de la magulladura durante el período de
almacenamiento. También, la identificación de los cambios en el
color (valor L*), o el progresivo oscurecimiento en el sector de yemas y tallos
después del pelado de los tubérculos, puede contribuir a entender
y cuantificar el fenómeno de magulladura. Esto además, puede ayudar
a desarrollar recomendaciones respecto del uso de las condiciones de almacenamiento,
ya que, de acuerdo a los resultados obtenidos la atmósfera de 5%CO2
6%O2 fue la más efectiva en reducir los cambios
en coloración (valores a* y b*), en tanto la AR y la con 5%CO2;
6%O2 mantuvieron la tonalidad y el grado de luminosidad
(L*) en los tubérculos magullados.
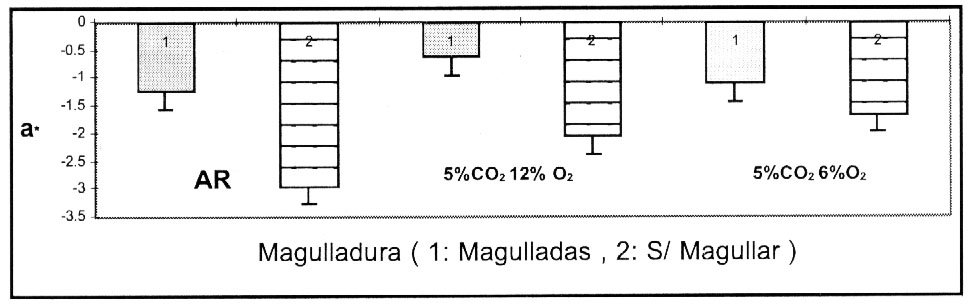
La figura 3, compara el efecto de la condición
de almacenamiento en el valor de a*. Las muestras se obtuvieron desde el interior
de la bodega de almacenamiento, bajo condiciones de iluminación intermitente,
lo que probablemente causó un grado de verdeamiento en los tubérculos,
los cuales fueron expuestos a la luz durante la toma de las muestras.
|
| |
Figura 3. Color verde (valor a*) de pulpa de papas cv
Ranger Russet, con y sin magulladuras , medido 2 meses después de
iniciado el almacenamiento en las 3 atmósferas. Almacenado a 6.6
ºC y 95 % HR.
Figure 3. Green coloration (a* value)
of flesh from potato cultivar Ranger Russet with and without bruising, determined
after 2 months of storage under 3 conditions. Storage at 6.6 ºC and
95% RH. |
Además, los tubérculos no magullados, exhibieron
un tono menos gris y aparentemente fueron menos afectados por la iluminación
de la sala, que aquellos magullados y mantenidos en las tres condiciones de
almacenamiento. El fenómeno de magulladura, causó un incremento
en el grado de tono gris, pero fue igualmente controlado por las atmósferas
de almacenamiento en 5% CO2 6%O2
y AR (Figura 3).
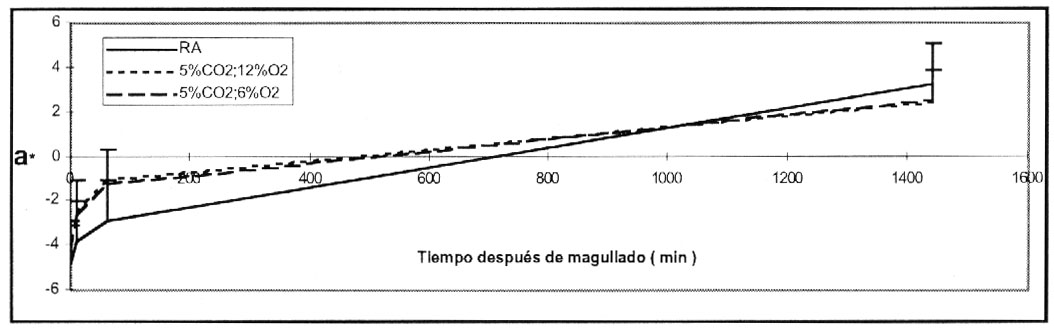
Tanto los tubérculos, magullados como los sin magullar,
mostraron una tendencia a incrementar el color verde, durante el tiempo de almacenamiento,
bajo las tres condiciones de atmósfera (Figura 4).
No se observaron diferencias significativas entre las dos condiciones de almacenamiento
en AC, en relación al valor de a*, obtenidos en tubérculos almacenados
en AR, que estuvieron en el mismo rango.
|
| |
Figura 4. Color verde (valor a*)
de pulpa de papa cv. Ranger Ruset almacenadas por 2 meses en las 3 atmósferas
, magulladas y evaluadas 24 h más tarde. Almacenadas a 6.6 º
C y 95 % R.R.
Figure 4. Green coloration (a* value) of flesh from potato
cultivar Ranger Russet stored for 2 months under 3 conditions. Storage
at 6.6 ºC and 95% RH. |
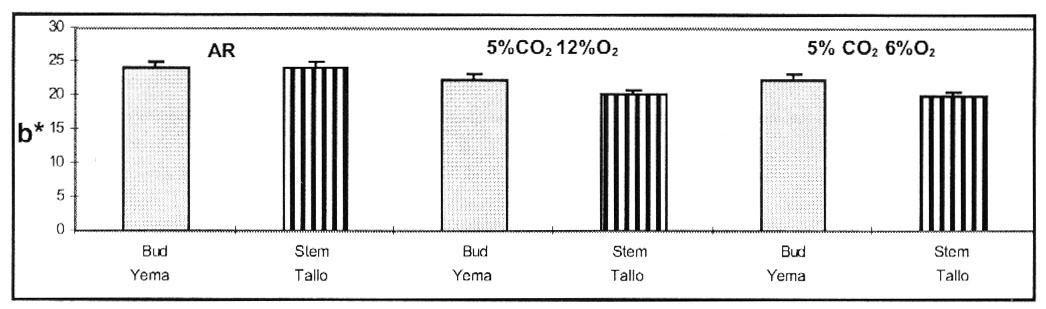
La figura 5, muestra que tubérculos
almacenados en AR, tuvieron un color amarillo más intenso (alto valor
b*) en los sectores del tallo y yema, respecto de los tubérculos mantenidos
en ambas AC. No se encontraron diferencias, entre las dos condiciones de AC,
pero el sector del tallo de los tubérculos tuvo valores menores de b*
que el de las yemas. El color amarillo en papas puede ser debido a los carotenoides,
pigmentación natural que actúa como antioxidantes (Francis,
1985). La sección de las yemas en los tubérculos, exhibió
una interacción no significativa entre Tiempo de Almacenamiento, Parte
del Tubérculo y Magulladura para el valor b*. La sección del tallo
de tubérculos magullados, mostró menor intensidad de color amarillo
que tubérculos no magullados (Figura 6).
|
| |
Figure 5. Color Amarillo (valor
b*) de porción de yemas y tallos de papas c.v. Ranger Russet, medido
2 meses despues de ingresar en las 3 atmósferas. Almacenadas a
6.6 º C y 95 % R.R.
Figure 5. Yellow coloration (b* value) of bud and stem
flesh from potato cultivar Ranger Russet determined after 2 months of
storage under 3 conditions. Storage at 6.6 ºC and 95% RH. |
| |
| |
|
|
| |
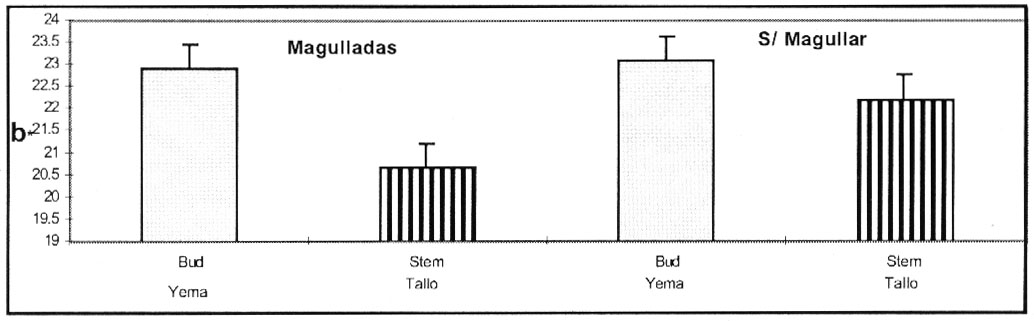
Figura 6. Color Amarillo ( valor
b*) de porción de tallos y yemas de pulpa de papas cv. Ranger Russet,
con o sin magulladuras, medido 2 meses despues de ingresar en las 3 atmósferas
. Almacenadas a 6.6 º C y 95 % H.R.
Figure 6. Yellow coloration (b* value) of bud and stem
flesh from potato cultivar Ranger Russet, with and without bruising, determined
after 2 months of storage under 3 conditions. Storage at 6.6 ºC and
95% RH. |
La diferente respuesta del sector de yemas y tallos, está
probablemente relacionada con su composición física y química.
La tirosina y la enzima PPO, responsable de la formación de melanina,
podrían estar localizadas en diferentes áreas de los tejidos,
tales como el tejido cortical del sector de tallos y tejido perimedular del
sector de las yemas (Craft, 1966). El efecto de magulladura,
causó cambios principalmente en el grado de luminosidad (valor L*) y
disminuyó la intensidad del color amarillo (valor b*) resultando en un
color oscuro.
Dwelle y Stallknecht (1976) establecieron,
que la modificación de la atmósfera con CO2
presurizado y N2, efectivamente bloquearon el sistema
de la enzima polifenol oxidasa, cuando el aire atmosférico se extrajo
desde la cámara a presión no quedando suficiente O2
disponible para que la reacción efectivamente ocurriera. En el presente
estudio, la combinación de CO2 y O2,
utilizada en las dos condiciones de AC a 6,6¥C
y 95% HR permitió obtener más información acerca de los
progresos del fenómeno de magulladura en papas cv. Ranger Russet, durante
el período de almacenamiento.
En condiciones de almacenaje, a temperaturas de 10¥C
o menores, utilizadas por Dwelle y Stallknecht (1976), el fenómeno
de magulladura necesita 24 a 48 h para su completo desarrollo. En este ensayo,
la temperatura de almacenamiento fue de 6,6¥C,
lo cual pudo haber causado la manifestación del color oscuro en tubérculos
magullados 24 h después de pelados.
CONCLUSIONES
Papas cv. Ranger Russet magulladas previo al almacenamiento,
exhibieron cambios significativos en la luminosidad (valor L*), pero no en los
colores verde (a*) y amarillo (b*). La zona apical de yemas y tallos, fue igualmente
susceptible al efecto de magulladura, según se reflejó de los
parámetros L*, a* y b*. Las papas magulladas mostraron un color oscuro
60 minutos después del pelado, y los valores de a* y b* estuvieron en
el mismo rango del gris.
Las magulladuras medidas en este estudio, no exhibieron una
decoloración a través de la pulpa, sino que sólo localmente.
De los resultados obtenidos, solo los valores de L* fueron más representativos
del daño por magulladura.
Las condiciones de AR y en 5%CO2
6%O2 fueron las más efectivas en prevenir el
oscurecimiento (valor L*) y con ello, evitar una disminución del valor
b* en la porción de las yemas de los tubérculos. La condición
de almacenamiento de 5%CO2 6%O2,
fue efectiva en reducir el grado de tono verde (valor a*) y también el
amarillo (valores de b*).
BIBLIOGRAFIA
Blight, D.P. ; Hamilton, A.J. 1974. Varietal
Susceptibility to Damage of Potatoes. Potato Res. 17 : 261-270.
Craft, C.C. 1966. Localization and Activity
of Phenolase in the Potato Tuber. Am. Potato J. 43 : 112-117.
Dwelle, R.B. ; Stallknecht, G.F. 1976. Rates
of Internal Blackspot Bruise Development in Potato Tubers Under Conditions of
Elevated Temperatures and Gas Pressures. Am. Potato J. 53 : 235-241.
Francis, F.J. 1985. Pigments and Other Colorants.
In : Owen R. Fennema (ed.). Food Chemistry. 2n ed. Revised and Expanded. Marcel
Dekker, INC. NY and Basel.
Gray, D. ; Hughes, J.C. 1978. Tuber Quality,
In The Potato Crop : The Scientific Basis for Improvement. Harris, P.M., Ed.,
Chapman and Hall, New York 504p.
Hudson, D.E. 1975. Relationship of Cell Size,
Intercellular Space and Specific Gravity to Bruise Depth in Potatoes. Am. Potato
J. 52 : 9-13.
Hunter Associates Laboratory. 1991. Hunter
Lab Associates’ Vision. Hunter Associates Laboratory, Inc., Reston, VA.
19p.
Johnson, L.F. 1972. A Simple Field Device
to Measure Potato Tuber Susceptibility to Bruise. Am. Potato J. 49 : 359-364.
Kadam, S.S.; Wankier, B.N.; Adsule, R.L. 1991.
Processing. In : D.K. Salunkhe, S.S. Kadam, and S.J. Jadhav (eds.). Potato :
Production, Processing, and Products. CRC Press, Inc., Boca Raton, FL.
Loyola, N. 1997. The Potential of Controlled
Atmosphere Storage for Fresh and Processing Potatoes. MS Thesis. Hort. and Land.
Arch. Dept. Washington State University.
Mathew, R. 1992. Impact Bruise Evaluation
of Potato. MS Thesis. Agric. Eng. Dept. Washington State University.
Mohsenin, N.N. 1986. Physical Properties
of Plant and Animal Materials. Gordon and Breach Science Publishers, NY.
Noble, R. 1985. The Relationship Between
Impact and Internal Bruising in Potato Tubers. J. Agric. Eng. Research 32 :
237-245.
Ott, L.R. 1992. An Introduction to Statistical
Method and Data Analysis. 4th ed. Marion Merrell Dow, Inc. Belmont, CA.
Pavek, J.J.; Corsini, D.L.; Love, S.L.; Hane,
D.C.; Holm, D.G.; Iritani, W.M.; James, S.R.; Martin, M.W.; Mosley, A.R.; Ojala,
J.C.; Stanger, C.E.; Thornton, R.E. 1992. Ranger Russet : A Long Russet Potato
Variety for Processing and Fresh Market with Improved Quality, Disease Resistance,
and Yield. Am. Potato J. 69 : 483-490.
SAS Institute Inc. 1991. SAS User’s
Guide : Statistics, Version 6.11 Ed., Cary, NC.
Sawyer, R.L. ; Collin, G.H. 1960. Blackspot
of Potatoes. Am. Potato J. 37 : 115-120.
Timm, H.; Yamaguchi, M.; Hughes, D.L.; Weaver,
M.L. 1976. Influence of Ethylene on Blackspot of Potato Tubers. Am. Potato J.
53 : 49-53.
USDA. 1990. Agricultural Statistics, United
States Government Printing Office, Washington DC.
USDA. 1994. Agricultural Statistics, United
States Government Printing Office, Washington DC.
Wendell, R. 1995. Measurement and Analysis
of Impact Failure in Potato Tuber Tissue. PhD Thesis. Agric. Eng. Dept. Washington
State University.
Zhang, F.; Britton, M.G.; Townsend, J.S.
1989. Potato Bruise Prediction by Finite Element Method. ASEA meeting presentation
paper n× 89 – 6056.