AGRO SUR 33 (2) 74-86 2005
CIENCIA AGRARIA
FRECUENCIA DE REPOSICION HIDRICA EN OLIVOS (Olea europaea L.) BAJO RIEGO POR GOTEO EN UN SUELO FRANCO ARCILLOSO
Frequency
of hydric reposition in olives (Olea
europaea L.) under drip irrigation in
a clay loam soil
Celerino Quezada L. 1, Alejandro Venegas V., Hernán García A. y Jorge Ocampo
Universidad
de Concepción, Facultad de Agronomía, Casilla 537, Chillán, Chile.
1 e-mail: equezada@udec.cl
ABSTRACT
A field experiment
was carried out in the 2002-2003 growing season in order to determine
the effect of irrigation frequency on the vegetative growth of two year
old olives (Olea europaea L.), cv. Arbequina, under drip irrigation
in a clay loam soil. The study was undertaken at the Agricultural Experimental
Station of the University of Concepción, in Chillán, Chile (36°34’
S lat, 72°06 W long). The treatments of irrigation frequencies were T1,
once daily; T2, once every 4 days; and T3, once every 7 days. The following
soil and vegetative parameters were determined volumetric soil water content,
trunk diameter, plant height, leaf area, shoot length, dry matter and
diffusive resistance. The results showed that in clay loam soils, low
irrigation frequency did not provoke statistically significant differences
in the vegetative behaviour, except in the plant height and leaf area
where a greater growth was observed with T1 and T2, respectively.
Low irrigation frequencies increased the wet bulb size and the root growth
that will provoke accumulative effects on the vegetative behaviour in
the following season. On the other hand, a high irrigation frequency produced
a superficial water concentration and a declining of diffusive resistance
Key words: porometer,
TDR, wet bulb, diffusive resistance, Arbequina
RESUMEN
En la temporada 2002-2003 se evaluó el efecto de la frecuencia de riego en el desarrollo vegetativo de olivos (Olea europaea L.) cv Arbequina de dos años de edad, regados por goteo en un suelo franco arcilloso. El ensayo se realizó en la Estación Experimental de la Universidad de Concepción (36°34’ lat. Sur; 72°06’ long Oeste, 144 m.s.n.m.), Chillán, Chile. Los tratamientos fueron T1, riego diario; T2, riego cada 4 días; y T3 riego cada 7 días. Se midieron parámetros de suelo, como contenido volumétrico de agua del suelo y parámetros vegetativos como diámetro de tronco altura de la planta, área foliar, largo de brote, materia seca y resistencia difusiva. Los resultados de solo una temporada de investigación demostraron que bajas frecuencias de riego no afectaron significativamente el desarrollo vegetativo del cultivo. Sólo la altura de la planta y el área foliar mostraron diferencias significativas (P£ 0,05), lográndose los mayores crecimientos con T1 y T2, respectivamente. La baja frecuencia de riego aumento las dimensiones del bulbo húmedo y el desarrollo radicular, lo que producirá efectos acumulativos en el comportamiento vegetativo de las temporadas siguientes. Por su parte, un riego con alta frecuencia (diana) genera una concentración de humedad en superficie y una disminución de la resistencia difusiva.
Palabras claves: porómetro, TDR, bulbo húmedo, resistencia difusiva Arbequina.
INTRODUCCION
El olivo (Olea europaea, L.) es una especie originaria del Asia Menor, desde donde se extendió a la cuenca del Mediterráneo, zona que concentra más del 90% de la superficie mundial (Guerrero, 1994). En el país, el olivo alcanza una superficie de 5.624 ha, distribuidas entre la I y IX Regiones (ODEPA, 2004) y concentradas en las Regiones de Atacama y Tarapacá, con 3.008 ha, que representan un 67% de la superficie olivícola nacional (INE, 1998). El olivo es una especie de gran interés en muchas áreas del mundo, no sólo por sus características agronómicas y económicas, sino que por su resistencia a la sequía y su capacidad para crecer en suelos de calidad pobre, siendo un rubro muy interesante para zonas áridas y semi-áridas (Fernández y Moreno, 1999).
La hoja del olivo presenta un color verde oscuro brillante con una gruesa cutícula protectora en el haz (Barrios, 2000) y por el envés presenta una coloración gris, debido a la presencia de tricomas peltados que le confieren la vellosidad característica (López, 1996). Los estomas están ubicados en el envés de la hoja, en ligeras depresiones donde se crea un microclima más húmedo, con una densidad entre 250 mm-2 a más de 700 mm-2, dependiendo del cultivar y del estado nutricional (Fernández y Moreno, 1999). Además, el poseer este tipo de tricoma, les otorga a las hojas la posibilidad de inmovilizar el aire cercano a los estomas y así aumentar la resistencia a la difusión del vapor de agua (Grammatikopoulus et al., 1994). El control estomático disminuye la transpiración cuando las condiciones de estrés hídrico y humedad relativa así lo requieran (Fernández et al., 1997; Rojas y Roldan, 1996). Sin embargo, la intensidad lumínica influye en la apertura total del poro, al igual que bajas concentraciones de CO2. Además, el potencial hídrico de las hojas ejerce control sobre la apertura y cierre estomático (Lira, 1994). Sin embargo, la conductancia estomática disminuye regularmente con el potencial hídrico, no existiendo un valor umbral de cierre estomático (Fernández et al., 1997; Castel y Fereres, 1982). Un bajo potencial hídrico xilemático le permite extraer agua desde el suelo cuando la humedad es baja, incluso si es inferior al punto de marchitez permanente (PMP). El alto módulo de elasticidad de sus células, también ayuda a extraer agua de suelos normalmente considerados secos (Dichio et al., 1997). Xiloyannis et al., (1999) señalan otras formas de adaptación a la sequía, como el mantenimiento de un alto turgor en las células de las raíces, evitando o retardando la separación de éstas de las partículas de suelo. Otro mecanismo es el ajuste osmótico, tolerando temporalmente largos períodos de déficit hídrico (Dichio et al., 2003).
Este cultivo responde muy bien a la aplicación de riego, favoreciendo el desarrollo vegetativo, frutal y nutricional de la especie (Ferreyra et al., 2001). El efecto del déficit hídrico sobre el crecimiento y producción del olivo se puede presentar como una reducción del crecimiento y número de flores al año siguiente, menor número de flores perfectas y pistiladas, floración incompleta, aumento del añerismo y aborto ovárico, menor tamaño del fruto, mayor contenido de potasio en los frutos, adelanto de la madurez y menor contenido de aceite en el fruto (Ferreyra et al., 2001). El desarrollo del sistema radicular se produce en las capas más superficiales del suelo y es muy sensible a la hipoxia, por lo cual los suelos de mal drenaje son inadecuados para el cultivo (Fernández y Moreno, 1999). El olivo tiene bajos requerimientos hídricos y presenta una alta eficiencia de uso del agua en relación a otros frutales, debido al mecanismo de control estomático sobre la transpiración y bajos coeficientes de cultivo, especialmente en los meses de mayor demanda atmosférica (Fernández y Moreno, 1999). El comportamiento de los parámetros de rendimiento en relación al régimen de riego determina que el completo reemplazo de la ETc no es recomendable y los mejores resultados se obtienen aplicando sólo el 66% de la ETc (d'Andria et al., 2004).
El sistema de riego localizado de alta frecuencia es el más adecuado, con un lateral por hilera y 4 a 6 goteros por árbol generando un bulbo húmedo de diferentes dimensiones, en función del caudal del emisor, tiempo de riego y textura del suelo. Sin embargo, los riegos de alta frecuencia se consideran mas apropiados para suelos de texturas medias a gruesas, ya que en suelos arcillosos y de mayor capacidad de retención de agua, se pueden generar condiciones de saturación y asfixia en la zona radicular (Selles et al., 2003) Por lo anterior, la presente investigación tuvo como objetivo evaluar el efecto de diferentes frecuencias de riego en el crecimiento vegetativo del olivo de segunda hoja, bajo riego por goteo en un suelo de textura franco arcillosa.
MATERIALES Y MÉTODOS
El ensayo se realizo en la VIII Región, en la Estación Experimental «El Nogal» de la Universidad de Concepción (36°34’ lat. Sur; 72°06’ long. Oeste, 144 m.s.n.m.) Chillán, Chile, durante la temporada comprendida entre noviembre de 2002 a marzo de 2003. El sitio experimental esta ubicado en el valle central regado con una precipitación histórica anual de 1055 mm concentrada en los meses de invierno Pertenece al agroclima Chillán, que se clasifica dentro de un clima mediterráneo temperado (Del Pozo y Del Canto 1999).
El suelo es de origen volcánico, profundo y de textura franco arcillosa, perteneciente a la Serie Diguillín (medial, thermic Humic Haploxerands). El pH varía entre 5,8 a 6,2. La capacidad de uso del suelo es IIs (Carrasco, 1998). Las características físicas del suelo se presentan en el Cuadro 1.
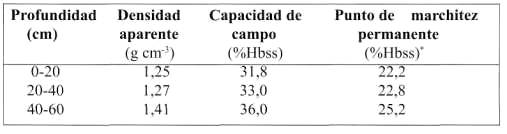 |
| (*) Humedad base suelo seco |
El marco de plantación fue de 1,5 metros sobre hilera y 4 metros entre hilera con una población de 1.666 pl ha-1. El cultivar correspondió a Arbequina de dos años, clon I -18 de vigor inferior a 20 - 30% a la Arbequina estándar (Casté 1999).
El ensayo se fertilizó con 50 g de nitrógeno por árbol en la forma de salitre sódico, parcializado en 9 oportunidades. El control de malezas se realizo periódicamente en la base del árbol en forma manual y entre hilera en forma mecanizada. La aplicación de los tratamientos se inicio el 30 de noviembre de 2002.
Se regó por goteo con un lateral por hilera y dos emisores por planta de 4 L h-1, ubicados a 20 cm del tronco. Antes de las aplicaciones se midió el coeficiente de uniformidad del sistema cuyo resultado fue 89% (López et al., 1997). Los datos de evaporación de bandeja se obtuvieron de la Estación Meteorológica de la Universidad de Concepción Campus Chillán.
El diseño utilizado correspondió a completamente al azar con tres tratamientos y tres repeticiones. Se contó con 9 hileras orientadas en dirección Norte-Sur y cada repetición estuvo formada por 5 árboles. El factor en estudio fue la frecuencia de negó con los siguientes tratamientos:
Tratamiento (T1).
Riego por goteo diario.
Tratamiento (T2). Riego por goteo cada 4 días.
Tratamiento (T3). Riego por goteo cada 7 días.
Estado hídrico del suelo. El contenido volumétrico de agua en el suelo se monitoreó cada 7 días utilizando un sensor dieléctrico ThetaProbe TDR (Delta T Devices, Cambridge, England), en las estratas 0-20, 20-40 y 40-60 cm de profundidad, a través de tubos de acceso de PVC.
Estado hídrico de las plantas. Se midió la resistencia difusiva, que es la resistencia que opone la hoja a la pérdida de vapor de agua a través de los estomas y la cutícula (Peretz et al., 1984), mediante un porómetro de difusión AP-4 (Delta T Devices, Cambridge, UK). Para ello se eligió un árbol por repetición y se evaluó una hoja sana ubicada en el tercio superior expuesta al sol con orientación oriente o poniente. Estas lecturas se efectuaron a las 07:00, 09:00, 14:00, 16:00, 19:00 horas y fueron realizadas cada 15 días durante toda la temporada.
Crecimiento de la temporada. Cada 15 días se midieron los siguientes parámetros en el total de los árboles del ensayo:
a)
Diámetro del tronco (mm), a 10 cm de la superficie del suelo, con un Vernier.
b) Largo
promedio de brotes (cm), con huincha métrica flexible de mano.
c) Área
foliar (cm2). Se seleccionaron al azar 3 hojas por árbol y
se utilizó el medidor de área foliar LI-COR ( EI-3000, USA).
d) Número
de hojas por planta. Se contabilizaron las hojas de cada árbol y se multiplicó
por el área foliar promedio para obtener el área foliar por repetición.
e) Altura
de la planta (cm), con huincha rígida de mano.
f) Números
de ramas.
g) Número
de brotes laterales.
h) Materia seca
(g). Al término de la temporada de riego se evaluó una planta representativa
por repetición, la cual se dividió en tres partes: raíces, tronco con
ramas y hojas. Las muestras fueron secadas a horno por 72 horas a 105°C,
luego se obtuvo la materia seca por órgano.
Análisis estadístico. Se utilizó el programa computacional SAS (SAS Institute Inc., 1999) y el test DMS (diferencias mínimas significativas) con un 95% de confianza. Los datos en porcentaje fueron normalizados a través de la transformación (x + 0,5)1/2 antes de ingresarlos al programa (Steel y Torrie, 1995).
RESULTADOS Y DISCUSIÓN
Agua aplicada.
La evapotranspiración se calculó de acuerdo a la fórmula: ETc = Eb x Kb x Kc , utilizando un Kb de (0,8) y un kc de (0,15). El valor que se asumió fue el 30% de un árbol adulto (Kc= 0,5), dado que no existe kc para olivos de tres años. Este coeficiente de cultivo es constante durante todo el año, ya que el desarrollo del árbol no pasa por las etapas de brotación y defoliación propias de los árboles frutales de hoja caduca (Goldhamer et al., 1994; Ferreyra et al., 2001).
El riego comenzó el 30 de noviembre 2002 y finalizó el 30 de marzo 2003. El volumen de agua aplicado en la temporada fue de 428 m3 ha-1 para todos los tratamientos, con un promedio por riego de 3,6 m3 ha-1 para T1; 14,3 m3 ha-1 para T2 y 25,2 m3 ha-1 para T3. Los datos se presentan en el Cuadro 2.

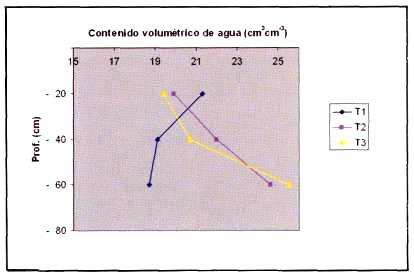
Estado hídrico del suelo. El contenido volumétrico de agua en función de la profundidad para la temporada se presenta en la Figura 1. Los contenidos para T1 fueron disminuyendo con la profundidad, siendo mayor en la primera estrata, con 21,3% y menor en la tercera con 18,7%. Esto puede ser explicado, considerando que el volumen de agua aplicado no permitió que el frente húmedo se distribuyera a mayor profundidad (Riquelme, 2003). Además, suelos de textura franco arcillosa tienen una alta retención de agua, debido a la mayor cantidad de microporos, lo que pudo causar que los aportes de agua se concentraran en los primeros 20 cm. En cambio en T2 y T3 hubo un aumento de la humedad en profundidad, con valores máximos de 24,6% y 25,5% respectivamente, ambos medidos en la tercera estrata. Los menores valores de T2 y T3 se presentaron en la primera estrata con 19,9% y 19,4% respectivamente. Esto se puede explicar porque al aplicar un mayor volumen de agua por riego el frente húmedo se distribuye a mayor profundidad, donde es menor la extracción por parte de la planta, ya que las raíces se concentran en los primeros 30 cm principalmente (Holzapfel et al., 2001; Ferreyra et al., 2001).
En el Cuadro 3 se aprecia el porcentaje de agotamiento de la humedad aprovechable por estrata y el promedio ponderado del perfil para cada tratamiento en contenido volumétrico. Estos resultados reflejan una distribución distinta de la humedad entre los tratamientos (T1 v/s T2 y T3), Se observa que en T1 el mayor agotamiento de la humedad ocurre a una profundidad de 20 a 60 cm con un porcentaje de agotamiento sobre el 66%, ya que por el bajo volumen de agua aplicado el bulbo húmedo no se distribuyó a una mayor profundidad. La humedad se concentró en los primeros 20 cm, con un porcentaje de agotamiento de 33,3%, debido principalmente a una frecuencia de reposición diaria. En promedio para el perfil T1 presenta un agotamiento del 55, 3%, respecto a los otros tratamientos, T2 con 40,1 % y T3 con 41,7%. Esto claramente está indicando una condición que para la planta puede ser inadecuada, y probablemente acumulativa, con efectos sobre los parámetros vegetativos en las temporadas siguientes.
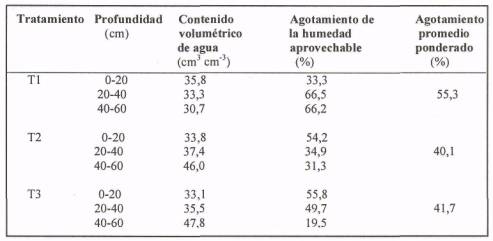
Para T2 el mayor agotamiento se presenta en la estrata de 0 a 20 cm por la mayor evaporación superficial y mayor densidad radicular en superficie, debido a que el bulbo húmedo alcanzó mayor profundidad por el mayor volumen de agua aplicado. El promedio ponderado fue de 42% que representa una condición más adecuada para la planta.
Para T3 el mayor porcentaje de agotamiento se presentó en los primeros 40 cm de profundidad. Al reponer el agua con una menor frecuencia, la evaporación es mayor, al igual que la extracción por parte de la planta, debido a las mayores dimensiones del bulbo húmedo; sin embargo, el porcentaje de agotamiento a mayor profundidad fue menor para ambos tratamientos, ya que el olivo es una planta que tiene un sistema radicular poco profundo, concentrando la mayor cantidad de raicillas en los primeros 30 cm, zona en la cual la absorción de agua es mayor. Acá, como se vio en la figura del perfil volumétrico de agua, existe una mayor distribución del agua en profundidad.
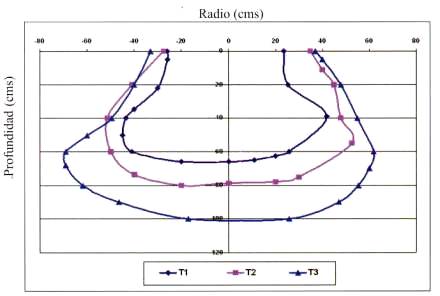
Bulbo húmedo. Se estableció un valor umbral equivalente al 25% de la humedad aprovechable para determinar el área del bulbo húmedo como condición adecuada de humedad para el desarrollo del cultivo. Se aprecia en la Figura 2 que T3 presenta el mayor tamaño de bulbo (0,94 m2) con más de 90 cm de profundidad y 132 cm de ancho. Luego T2 (0,68 m2), con 80 cm de profundidad y 104 cm de ancho. El bulbo de menor tamaño fue T1 con (0,43 m2) presentando 66 cm de profundidad y 87 cm de ancho. Estos resultados concuerdan con los encontrados por Sellés et al. (2003), Holzapfel et al. (1991) y Riquelme (2003) quienes en estudios realizados en vid, almendro y cerezo respectivamente, encontraron que menores frecuencias y mayores volúmenes de agua por riego generan un aumento del tamaño del bulbo húmedo.
En el Cuadro 4 se observa que el crecimiento horizontal de raíces fue mayor en T3 con 44,4 cm, luego le sigue T1 con 41,2 cm mientras que T2 alcanzó los 35,4 cm; sin embargo, no hubo diferencias significativas (P>0,05). En cuanto a la profundidad radicular, fue mayor en T3 y T2 con 69,9 cm y 65,1 cm respectivamente, mientras que T1 alcanzó una profundidad de 62,3 cm, no apreciándose diferencias significativas (P>0,05). El hecho que T3 tenga un mayor desarrollo de raíces, tanto en crecimiento horizontal como vertical, se debe a que el bulbo húmedo fue de mayor volumen, de tal manera que permitió condiciones favorables para el desarrollo radicular producto de una buena relación aire-agua que evitó condiciones de asfixia. En T2, la profundidad radicular también es alta debido a las mismas razones anteriormente señaladas. Entre los principales factores que inciden en el desarrollo radicular, están la disponibilidad de agua y la resistencia mecánica del suelo (Kramer, 1983).
El crecimiento lateral de T1 fue mayor que T2, lo que se puede deber al hecho que en T1, con reposición diaria, se pueden generar condiciones de anaerobiosis lo que hace que aumente el desarrollo de raíces laterales (Gil, 1997).
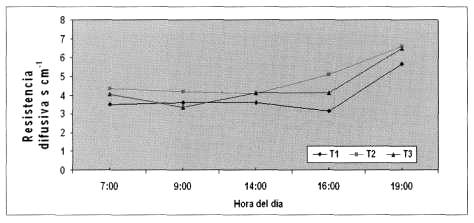
Resistencia difusiva o estomática. Los resultados para la temporada se presentan en la Figura 3. La mayor resistencia difusiva se determinó a las 19:00 horas, ya que a esa hora la mayor parte de los estomas se encuentran cerrados, con valores de 5,65 s cm-1 para T1; 6,6 s cm-1 para T2 y 6,5 s cm-1 para T3. Estos resultados concuerdan con los obtenidos por Punthakey et al. (1984), Dinamarca (2001) y Riquelme (2003), quienes encontraron valores más altos de resistencia difusiva al atardecer. En cambio los menores valores se midieron a las 9:00 horas, lo que coincide con lo reportado por Fernández et al. (1997) y Giménez et al. (1997), quienes encontraron estomas completamente abiertos a las 10:00 horas, cuando el déficit de presión de vapor es bajo, al igual que la transpiración. Holzapfel et al. (2001), evaluaron distintos niveles de agua en naranjo y encontraron valores de resistencia estomática entre 4 y 5 s cm-1 para plantas con un adecuado suministro hídrico, similar a los resultados de esta investigación. Por su parte, Flores (2005) determinó valores de resistencia estomática entre 0,14 a 2,06 s cm-1 en uva de mesa con diferentes tratamientos de aplicación de agua, entre 40% a 120% del volumen de agua requerido. Concluyó que los valores son muy variables a lo largo de la temporada de medición, por lo que no sería un buen índice para representar el estado hídrico de las plantas y su medición no sería tan relevante como el potencial hídrico xilemático.
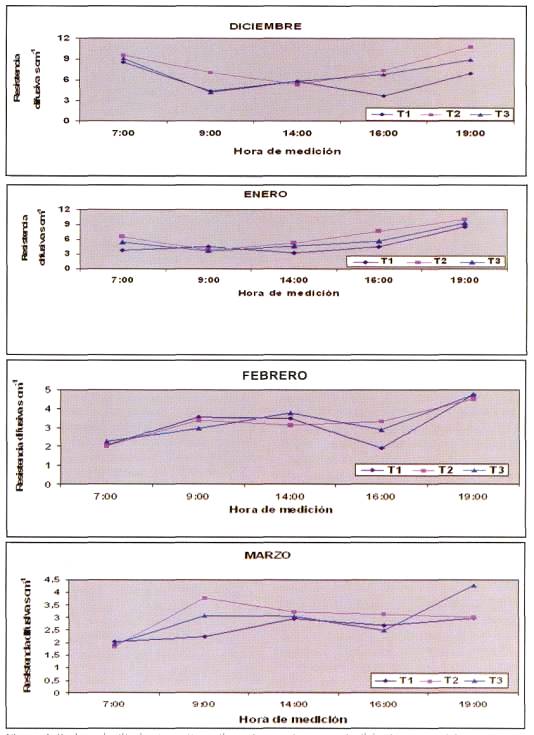
La Figura 4 presenta la resistencia difusiva en 4 momentos distintos de la temporada. El comportamiento variable durante la temporada de riego permite concluir que la planta retiene un poco más el agua en los meses de diciembre y enero, en comparación con los meses de febrero y marzo. Esto se explica considerando que la demanda evaporativa de la atmósfera es mayor en los primeros meses y la humedad relativa es menor (Giménez et al, 1997; Rojas y Roldan, 1996). Resultados similares fueron obtenidos por Dinamarca (2001), quien encontró en naranjo un aumento de la resistencia difusiva en los primeros meses de riego. El hecho que las plantas en T1 tengan menor resistencia difusiva durante el día indica que con este tratamiento la planta transpira más, tiene una mayor cantidad de estomas abiertos y además es mayor la fotosíntesis, por ende debiera producir mayor cantidad de materia seca. Sin embargo, en T2 y T3 la resistencia difusiva fue mayor, lo que podría indicar un estado de estrés por parte de la planta, debido a los mayores contenidos de agua de estos tratamientos, ya que la planta es sensible a la falta de oxígeno en las raíces, lo que aumentaría la producción de ácido abscísico, el cual provoca el cierre de los estomas (Lira, 1994).
Parámetros vegetativos. De acuerdo a los resultados que se presentan en el Cuadro 5, los únicos parámetros vegetativos que presentaron respuesta a la aplicación de diferentes frecuencias de riego fueron altura de la planta y área foliar.
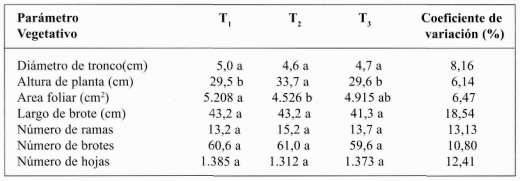
Altura de la planta. Presentó diferencias significativas (P≤0,05) entre T2 y T1 al igual que entre T2 y T3. Sin embargo, no existieron diferencias significativas entre T3 y T1 (Cuadro 5). T2 es el que genera la mayor altura con 33,7 cm; le sigue T3 con 29,6 cm; y luego T1 con 29,5 cm. El tratamiento T2 entre los 20 y 40 cm de profundidad de suelo, tiene el mayor contenido de agua, con un 65,1% de la humedad aprovechable y además coincide con la profundidad efectiva de raíces, por lo tanto la planta absorbe mejor los nutrientes, ya que la aireación del suelo y el porcentaje de agua permitieron una mayor exploración radicular, incidiendo positivamente en el crecimiento de los árboles (Coleto, 1989).
Materia seca
En el Cuadro 6 se presentan los resultados de la materia seca acumulada obtenida por hoja, raíz y rama de la planta, expresados en gramos (g) y porcentajes (%) de cada parte en relación al árbol completo. Estos parámetros no presentaron diferencias significativas entre los tratamientos (P>0,05).
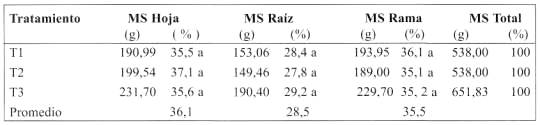
Porcentaje materia seca de raíz. El porcentaje de raíces se expresó como relación entre peso seco de raíces y peso seco total del árbol extraído. T3 presenta el mayor porcentaje de raíces con 29,2%; luego T1 con 28,5% y T2 con 27,8%.
Según Sierra (2003), el porcentaje de raíces de un árbol es considerado aceptable si está sobre un 20 a 25% de la materia seca total. Por lo tanto, la acumulación de MS de la raíz se consideró adecuada. Según Coleto (1989) el crecimiento de la raíz depende del suministro de hidratos de carbono y de reguladores de crecimiento sintetizados en la parte aérea. Así, todos los factores que afectan a la fotosíntesis y la traslocación de productos de los brotes a las raíces inciden en el crecimiento de la raíz. Las reservas acumuladas en la planta son más importantes a partir del segundo año de vida, mientras que durante el primero, el crecimiento radicular depende más de los productos fotosintetizados. Es por esto, que el aumento del área foliar estimula el crecimiento de la raíz.
Porcentaje materia seca de hojas. El valor más alto se encontró en T2 con 37,1%, seguido de T3 con 35,6% y por último T1 con 35,50% (Cuadro 6). Llama la atención que T2, si bien presentó la menor área foliar, fue el que generó mayor materia seca, lo que tendría explicación en la teoría funcional de Brower (1983). Ésta sugiere que la distribución de los recursos entre brotes y raíces es determinada dinámicamente por la disponibilidad de los recursos en sus respectivos ambientes (atmósfera y suelo). Si los recursos edáficos son abundantes, una mayor proporción de hidratos de carbono serán enviados hacia los brotes, resultando un incremento del crecimiento de la parte aérea de la planta con un consecuente aumento en la capacidad fotosintética. Sin embargo, cuando estos recursos son escasos, una cantidad excesiva de hidratos de carbono serán acumulados en los brotes y no generaran crecimiento debido a la escasez de minerales. En este caso los hidratos de carbono serán transportados a la raíz, incrementando su tamaño y mejorando el suministro de minerales. Esta teoría sugiere que las condiciones ambientales son la principal determinante de la relación raíz/brote
Porcentaje materia seca de ramas. El valor más alto se encontró en T1 con 36,1%; luego T3 con 35,2% y por último T3 con 35,1%. Según Coleto (1989), esto tendría explicación en la parte radicular, ya que esta provee a la canopia con agua, minerales, nutrimentos orgánicos, citoquinina, giberelina, ácido abscísico, todos de gran impacto en su desarrollo. Estos porcentajes coinciden con el área foliar que fue mayor en T1 y menor en T2, lo que también tendría explicación de acuerdo a la teoría funcional de Brower (1983).
CONCLUSIONES
- En olivos de dos años de edad establecidos en suelos de textura franco arcillosa, bajas frecuencias de riego no afectaron significativamente el desarrollo vegetativo del cultivo.
- La frecuencia de riego solo tuvo efecto en la altura de la planta y el área foliar, lográndose los mayores crecimientos con riegos cada 4 días y riegos diarios, respectivamente.
- La aplicación de agua mediante el sistema de goteo con menor frecuencia genera bulbos húmedos de mayores dimensiones y mayor desarrollo radicular, lo que generara efectos acumulativos en los parámetros vegetativos en las temporadas siguientes.
- Por su parte, un riego con alta frecuencia (diaria) genera una concentración de humedad en superficie y una disminución de la resistencia difusiva.
BIBLIOGRAFIA
BARRIOS, C.O. 2000. El olivo emerge como buen negocio. Nuestra Tierra (207): 21-24.
BROWER, R. 1983.
Functional equilibrium: sense or nonsense. Neth. J. Agric. Sci. 31(4):
335-348
CARRASCO, P. 1998. Descripciones de suelos VIII Región. Universidad de Concepción, Facultad de Agronomía, Departamento de Suelo. Chillán. Chile. 75 p.
CASTÉ A. 1999. Producción de aceite de oliva en España e Italia. Agroeconómico (50): 8.
CASTEL J.; FERERES,
E., 1982. Responses of young almond trees to two drought periods in the
field. J. Hortic. Sci. 57(2): 175 187
COLETO, M.J.M. 1989. Crecimiento y desarrollo de las especies frutales (2a ed) Mundi-Prensa. Madrid, España. 140 p.
D’ANDRIA, R.; LAVINI, A.; MORELLI, G.; PATUMI, M.; TERENZIANI, S.; CALANDRELLI, D.; FRAGNITO, F., 2004. Effects of water regimes on five pickling and double aptitude olive cultivars (Olea europaea L.). Journal of Horticultural Science & Biotechnology 79(1): 18-25.
DEL POZO, A.; DEL CANTO, P. 1999. Áreas agroclimáticas y sistemas productivos en la VII y VIII Regiones. Serie Quilamapu N° 113. Instituto de Investigaciones Agropecuarias, Centro Regional Quilamapu. Chillán, Chile. pp. 53-54.
DICHIO, B.; NUZZO,
N.; XILOYANNIS, C.; CELANO, G.; ANGELOPOULOS, K. 1997. Drought stress-induced
variation of pressure volume relationships in Olea europea L.
cv. «Corantina» In: K. S. Chartzoulakis (Ed). Proceedings
of the 2nd Int. Symposium on Irrigation of Hort. Crops,
September 9-13, 1996, Chania, (449): 401-409
DICHIO, B.; ROMANO,
M.; NUZZO, N.; XILOYANNIS, C. 2002. Soil water availability and relationship
between canopy and roots in young olive trees (cv. Corantina) In:
C. Vitagliano and G.P. Martelli (Eds). Proceedings of the 4th
Int. Symposium on Olive Growing, September 25-30, 2000, Valenzano, Italia.
Acta Hortic. (586): 225-258
DICHIO B.; C. XILOYANNIS,
C.; ANGELOPOULOS, K.; NUZZO, V.; BUFO, S.; CELANO, G. 2003. Drougth-induced
variations of water relations parameters in Olea europaea In:
H. Lambers (Ed). Symposium on Dryland
DINAMARCA, I.O. 2001. Efecto del volumen de agua aplicado y fertilización sobre la producción de naranjos (Citrus sinensis) cv. Thompson de quince años. Tesis Ingeniero Agrónomo, Chillán, Chile Universidad de Concepción, Facultad de Agronomía. 28 p.
FERNANDEZ, J. E.;
MORENO, F. 1999. Water use by the olive tree. p. 101-162. In: M.
B. Kirkham (Ed.). Water Use and Crop Production. Food and Press Production,
N.Y., USA
FERNÁNDEZ, J.; MORENO,
F.; GIRÓN, I.; BLÁZQUEZ, O. 1997. Stomatal control of water use
in olives trees leaves. Plant Soil 190(2): 179-192
FERREYRA E.R.; SELLES, G.; SELLES, I. 2001. Riego deficitario controlado en olivos. Estrategias de riego para enfrentar situaciones de escasez de agua en frutales Boletín INIA N° 59. INIA, La Platina Santiago, Chile.
FLORES, F.A. 2005. Efecto de la aplicación diferencial de agua y fertilización en la producción de uva de mesa bajo riego por goteo: primer año. Tesis Ingeniero Civil Agrícola, Chillán, Chile, Universidad de Concepción, Facultad de Ingeniería Agrícola, 57 p.
GIL, S.G. 1997. Fruticultura: el potencial productivo crecimiento vegetativo y diseño de huertos de viñedos. Fac. Agron. (Ed). Universidad Católica. Santiago, Chile. 342 p.
GIMÉNEZ ,C.; FERERES,
E.; RUZ, C.; ORGAZ, F. 1997. Water relations and gas exchange of olive
trees: Diurnal and seasonal patterns of leaf water potential, photosynthesis
andstomatal conductance. In: K. S. Chartzoulakis (Ed). Proceedings
of the 2nd Int. Symposium on Irrigation of Hort. Crops. September
9-13, 1996, Chania, Crete, Greece. Acta Hortic. (449): 411-415
GOLDHAMER D.A.;
DUNAI, J.; FERGUSON, L.F., 1994. Irrigation requirements of olive
trees and responses to sustained deficit irrigation. In: S. Lavee
and I. Klein (Eds). 2nd Int. Symposium on Olive Growing, September
6-10, 1993, Jerusalem, Israel. Acta Hortic. (356): 172-175
GRAMMATIKOPOULUS,G.,
KARABOUR-NIOTIS, G.; KYPARISSIS, A.; PETROPOULOU,Y.; MANETAS, Y. 1994.
Leaf hairs of olive (Olea europaea) prevent stomatal closure by
ultraviolet-B radiation. Australian J
GUERRERO, A. 1994. Nueva olivicultura (3ª ed.). Mundi-Prensa. Madrid, España. 273 p.
HOLZAPFEL, E.A.;
FERERES, E.; VALENZUELA, A. 1991. Patterns of soil water distribution
and uptake by young almond trees under drip irrigation. Agro-Ciencia.
7(1): 39-46
HOLZAPFEL, E.A.; LÓPEZ, C.; JOUBLAN, J.P.; MATTA, R. 2001. Efecto del nivel de agua aplicada en la producción y desarrollo de naranjos cv. Thompson navel. Agro Ciencia. 17(1): 69-78.
INE (CHILE). 1997. VI Censo Nacional Agropecuario. Instituto Nacional de Estadísticas. Santiago, Chile. 443 p.
KRAMER, P.J. 1983.
Water relations of plants. Academic Press. New York, U.S.A. 489 p
LIRA, R.H. 1994. Fisiología vegetal. Trillas. México. 237 p.
LÓPEZ F., C. 1996. El cultivo del olivo. El campesino 127(4): 50-59.
LÓPEZ, J.R.; HERNÁNDEZ, J.M.; PÉREZ, A.; GONZÁLEZ, J.F. 1997. Frecuencia de aplicación. Riego a pulsos Pp. 179-182. In: Riego localizado (2da ed.) Mundi-Prensa, Madrid, España. ODEPA (CHILE). 2004. Estadísticas macrosectoriales y productivas (On line) www.odepa.gob.cl (23-01-2004).
PERETZ, J.; EVANS,
R.; PROEBSTING, E. 1984. Leaf water potential for management of high frequency
irrigation in apples. Trans. ASAE 27(2): 437-442
PUNTHAKEY, J.F.;
MCFARLAND, M.J.; WORTHIGTON, J.W. 1984. Stomatal responses to leaf water
potentials of drip irrigated peach (Prunus persica). Trans. ASAE.
27(5): 1442-1450
RIQUELME, F. 2003. Frecuencia de reposición de agua en cerezo (Prunus avium L.) bajo riego por goteo en un suelo franco arcilloso. Tesis Ingeniero Agrónomo, Chillán, Chile. Universidad de Concepción, Facultad de Agronomía. 30 p.
ROJAS, R.; ROLDAN,
J. 1996. Evapotranspiration mapping and irrigation scheduling for Olive
Trees. Pp. 449-454. In: C.R. Camp, E.J. Sadler, R.E. Yoder (Eds).
Evapotranspiration and Irrigation Scheduling. A.S.A.E. San Antonio, Texas,
U.S.A
SAS INSTITUTE. 1999.
SAS/STAT user’s GUIDE: statistics, Versión 6. 4th ed.
SAS Institute, Cary, North Carolina, USA
SIERRA, B., C. 2003. Antecedentes generales de factores críticos para el crecimiento aéreo y radicular de la vid. Micronutrientes y dinámica de los nutrientes solubles. 31 p. In: Curso Diagnóstico de la fertilidad, fertirrigación y nutrición de plantas con énfasis en vides, en suelos de zonas áridas. 26 al 28 de Mayo. INIA, Intihuasi. La Serena, Chile.
STEEL, R.G.D.; TORRIS, J.H. 1995. Bioestadística: principios y procedimientos. 2a Edición. México, D.F. McGraw-Hill. 622 p.
XILOYANNIS C.; DICHIO, B.; NUZZO, G.; CELANO, G. 1999. Defense estrategies of olive against water stress. In: I.T. Metzidakis and D.G. Voyiatzis (Eds). Proceedings of the 3rd Int. Symposium on Olive Growing, September 22-26, 1997. Chania, Crete, Greece. Acta Hortic. (474): 423-426.
Recepción de originales: 30 de marzo 2006