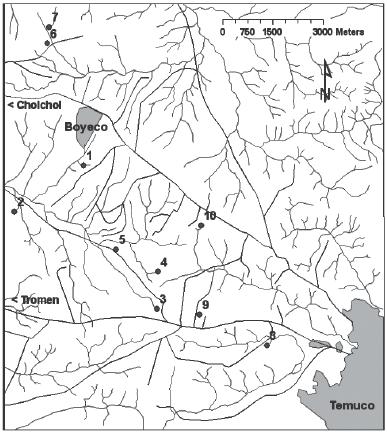
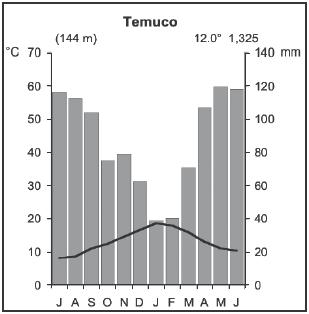
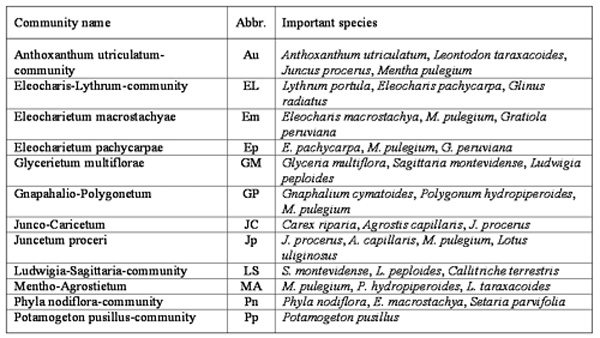
Ecology of the sampled plant species
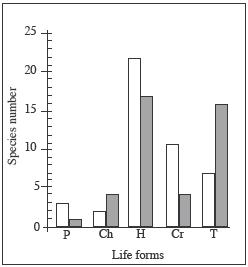
The biological spectrum (Figure 5)
corresponds to the therophitic phytoclimate typical of the northeast
region of Central Chile (San Martín y Ramírez 1986). In the studied
wetlands this biological spectrum was more developed in the dry summer
months (during the terrestrial phase sensu Pott y Remy 2000). In the
wettest season (winter and spring) perennial life-forms of hemicryptophytes,
hydrophytes and helophyes (cryptophytes) become dominant. The therophytes
were only found in the terrestrial phase, whereas the hemicryptophytes
and cryptophytes were found throughout the year. A similar seasonality
was found in Southern Chilean grasslands by Ramírez (1984).
 |
| |
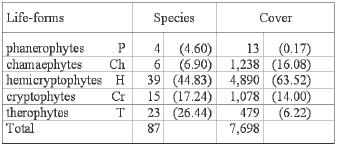
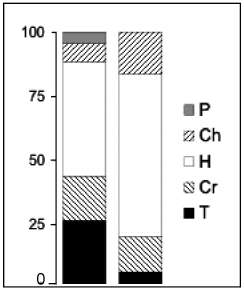
Figure
5. Life-form spectra according to the species number (left) and
the cover (right). Life-forms: phanerophyte (P), chamaephyte (Ch),
hemicryptophyte (H), cryptophyte (Cr) and therophyte (T).
Figura 5. espectro biológico de acuerdo al número de especies
(izquierda) y a su cobertura (derecha). Formas de vida: fanerófita
(P), caméfita (Ch), hemicriptófita (H), criptófita (Cr) y terófita
(T). |
Comparing the species list of this work with those
of Ramírez et al. (1983), 9 species of the native flora and 12
of the introduced flora have an affinity with the myrtaceous swamp forest,
which originally grew in these wetlands. On the other hand, typical
ephemeral wetland species are not very important, taking as a reference
the works of Ramírez et al. (1994), Bliss et al. (1998)
and Alvarez (2008). About 49 % of the total sampled species (43 species)
were terrestrial plants belonging to dry locations (Table
6). Swamp plants comprised 41 % (36 species) of all collected plants.
Aquatic plants accounted for only 9 % (8 species) of the flora, of which
5 are submerged plants (Myriophyllum aquaticum, Utricularia gibba,
Limosella australis, Crassula peduncularis, and Potamogeton pusillus)
and 2 were floating leaved species (Ludwigia peploides, Callitriche
palustris and Callitri-che autumnalis). These water plants
(almost the aerial parts) disappear in the dry seasons. Using cover
values as a measure, helophytes accounted for 70 % of the vegetation
and 50 % according to the total species number. This confirms the importance
of the swamp flora along the wetland shores and the seasonality of these
habitats, which represent amphibious ecosystems alternating very wet
periods with very dry ones (Deil 2005, Chambers et al., 2008).
Table
6. Distribution of plant species according to their habitat.
Cuadro 6. Distribución de las especies vegetales de acuerdo a
su hábitat. |
| |
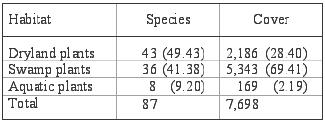 |
| Percentage
in parentheses |
Of the 87 species that formed the flora of the studied
wetlands, only 9 (10.34 %) were obligate ephemeral wetland plants (Zedler
1990). Eight were dicotyledons and one (Juncus stipulatus) was
a monocotyledon, all of which were native species, the hemicryptophytes
Centipeda elatinoides, Eryngium humifusum, Eryngium peudojunceum
and Phyla nodifora, the therophytes Glynus radiatus, Gnaphalium
cymatoides, Navarretia involucrata and Juncus stipulatus, and
the cryptophyte Lythrum portula. The majority of these are small
and colonize the central
zone of the wetlands which are dry in summer. The most important was
Phyla nodiflora, which was present in 9 wetlands and had an importance
value of 18.38. The next highest in importance were Gnaphalium cymatoides
(7 wetlands, importance value 14.38) and Centipeda elatinoides (8
wetlands, importance value 12.36). It is possible that these ephemeral
wetland specialists are dispersed from the native micro vernal pools
present in the neighbourhood and described by Ramírez et al., (1994).
The vast majority of the wetland flora were dry land
plants or wetland generalists (in both cases 39 species, corresponding
to 44.8 % of the total species number).
Plant diversity of the studied wetlands
A first analysis of the plant species diversity of
the wetlands can be made with the species number, also called alpha
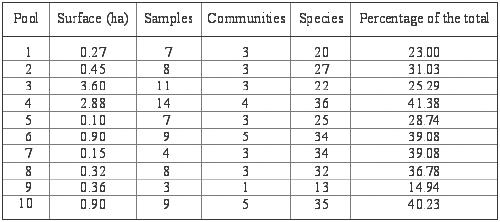
diversity (Whittaker 1972). The wetland number 9 had the lowest species
number (13 species, see also Table 7), whilst the
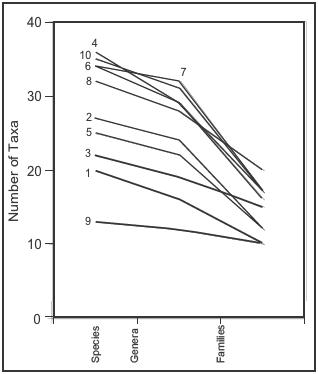
number 4 had the highest alpha diversity (36 species). However, the
relative diversity did not remain constant when considering the taxa
at different levels (genera and families, Figure 6).
Table
7. Area, number of vegetation samples, communities and species
in the studied wetlands.
Cuadro 7. Area, número de censos, comunidades vegetales y especies
en las lagunas estudiadas. |
| |
 |
 |
| |
Figure
6. Taxonomic diversity profile of the studied wetlands.
Figura 6. Perfiles de diversidad taxonómica de las lagunas estudiadas. |
Comparing the floristic composition between wetlands
(beta diversity sensu Whittaker 1972), we found a high degree of dissimilarity.
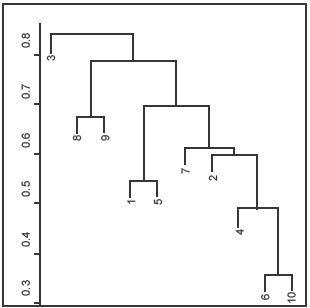
In the dendrogram resulting from the clustering analysis (Figure
7) we obtained 3 clusters at a distance of about 0.7; the first
cluster consists of wetland 3 only, the second wetlands 8 and 9, and
the third consists of the rest of the wetlands. However the last cluster
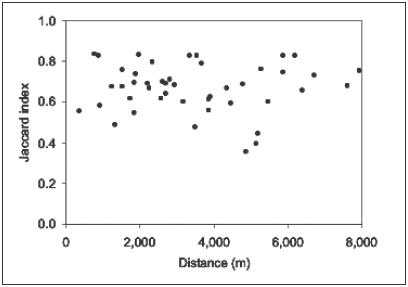
shows a high degree of heterogeneity. We found no correlation between
the distance between wetlands and their similarity (Figure
8).
|
| |
Figure
7. Dendrogram of the agglomerative hierarchical cluster analysis
using complete linkage and the Jaccard-index as a similarity measure.
Figura 7. Dendrograma de la clasificación jerárquica usando el
algoritmo "complete linkage" y el índice de Jaccard como medida
de similitud. |
|
| |
Figure
8. Floristic similarity between wetlands (pairwise comparison
by Jaccard-index) versus distance between compared wetlands.
Figura 8. Similitud florística entre las lagunas (comparaciones
pareadas usando el índice de Jaccard) en función a la distancia
entre los humedales comparados. |
All of the regression models of the species number
versus wetland area and number of present communities following the
power model (Arrhenius 1921) were not significant, including the single
models of wetland area (R2 = 0.029, p > 0.05) and number of communities
(R2 = 0.322,
p > 0.05) as well as the model including both parameters (R2 = 0.384,
p > 0.05).
Ordination analysis
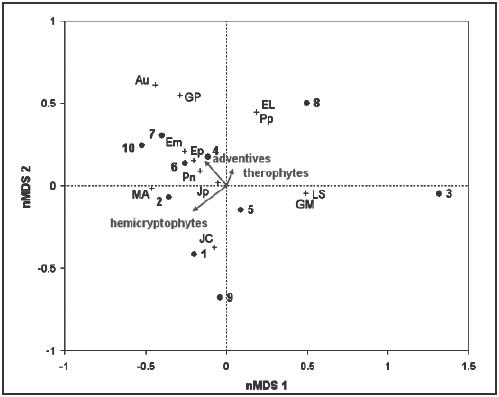
The wetlands in the ordination diagram (Figure
9) showed a high degree of dissimilarity according to their distribution.
Extreme cases were represented by the wetlands number 3 and 8 that lie
more separated from the rest of the wetlands. The distribution of the
plant communities in the diagram also showed a high level of dispersion.
|
| |
Figure
9. Ordination diagram of the temporary wetlands (o) using a non-metric
multidimensional scaling (nMDS) and the Bray-Curtis-index of dissimilarity.
Plant communities (+) and proportion of indicator plants (represented
as vectors) are added through post-hoc correlations. stress =
10.10. Abbreviation of the plant communities are shown in Table
1.
Figura 9. Ordenación de las lagunas temporales (o) usando el método
"non-metric multidimensional scaling" (nMDS) y el índice de disimilitud
de Bray-Curtis. Las comunidades vegetales (+) así como también
la proporción de plantas indicadoras (representadas por vectores)
fueron agregadas usando correlaciones post-hoc. stress = 10,10.
Las abreviaciones de las comunidades vegetales se presentan en
el Cuadro 1. |
The proportion of therophyte, hemicryptophyte and adventive
species were shown as vectors. The proportion of hemicryptophytes (see
also Table 8) had the highest correlation with the
ordination of the wetlands, whereas the lowest correlation was shown
by the therophytes. Therophytes and hemicryptophytes were negatively
correlated with each other; and they were poorly correlated with introduced
species.
Table
8. Correlation coefficients between the ordination axes and the
proportion of therophytes, hemi-cryptophytes and introduced plants
in the vegetation and for the presence of plant communities in
the studied wetlands.
Cuadro 8. Coeficientes de correlación entre las dimensiones
de ordenación y la proporción de terófitas, hemicriptófitas y
adventivas en la vegetación y la presencia de comunidades vegetales
en las lagunas muestreadas. |
| |
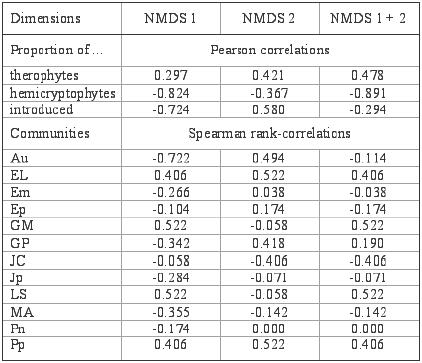 |
Community abbreviations are shown in Table
1. |
DISCUSSION
Wetland area and connectivity in the determination
of species richness
As has been shown to be the case for Chilean grasslands
(Oberdorfer 1960) and like Californian vernal pools (Barbour et al.,
2005), the wetlands near Temuco were floristically poor. They reached
a mean species density of only 8.4 species in 1,000 m2. Nevertheless,
the area of the wetlands did not play an important role in the determination
of the species number. Normally an analysis of the species-area relationship
on a small scale results in low correlations, a phenomenon known as
the small island effect (Lomolino y Weiser 2001). However, Alvarez (2008)
reported a weak but significant correlation between species richness
and area in smaller vernal pools (about 1.5 m2) near Temuco.
The fragmentation of this kind of habitat and the strong
contrast of the environmental conditions with the surrounding upland
(Holland y Jain 1981, Deil 2005) could result in a barrier for the dispersal
between ponds. Moreover, seasonal wetland species normally have adaptations
for restricted dispersal (Zedler 1990, Alvarez 2008). Under the scenario
of restricted dispersal of species between wetlands, a higher floristic
similarity was expected between wetlands closer together than between
those further apart. This hypothesis was rejected as a result of the
lack of correlation between distance and the similarity of the wetlands
(Figure 8). An accidental dispersal of diaspores by
water birds and livestock (diszoochory sensu Dansereau y Lems 1957)
could produce a broad dispersal of the wetland species throughout the
whole study area. Dispersal by epizoochorous mechanisms is not always
restricted to diaspores with morphological adaptations such as hooks
or viscous appendixes, as demonstrated by Ramírez et al., (2003)
in Southern Chilean grasslands.
Within wetland diversity
As a part of this study, two aspects of the so called
within wetland diversity were analysed: 1) The change in diversity over
different taxonomic levels (species, genera and families; Feoli y Scimone
1984). 2) The vegetation mosaics present in each wetland (Barbour et
al., 2003).
In the first case, different tendencies in the change
of diversity across taxonomic levels were found. For example, wetland
number 8 was the richest wetland at the family level (Figure
6), but not at the species level. Only wetland number 9 remained
the poorest at all taxonomical levels.
In the second case, the presence of different communities
within the wetlands was very variable (Table 7). Surprisingly
the number of communities in the wetlands was not correlated with the
species number. This variability was not only reflected in the number
of communities but by the community composition of each wetland (details
published by San Martín et al., 1998). Similar results have been
obtained analysing Californian vernal pools at a larger geographic scale
(Barbour≠ 2003).
Disturbance effects
Because all of the studied wetlands are anthropogenic
in origin, all are the result of disturbance (deforestation). In addition,
the seasonality in the environmental conditions that oscillate between
the extremes of total drought and flooding could be interpreted as a
natural disturbance, as it implies the frequent death of almost all
of the aereal part of the vegetation (Deil 2005). A continual disturbance
is provoked by the use of these wetlands as watering and grazing places.
This factor could produce effects through grazing, trampling and eutrophication
as a result of waste deposition in the wetlands. Manure transported
from the surrounding cultivated areas has an additional effect that
contributes to their eutrophication.
The high number of introduced species and their high
frequency around the wetlands (e. g. Mentha pulegium, Leontodon taraxacoides
and Polygonum persicaria; see also Table 3)
indicate both the secondary anthropogenic origin and the strong disturbance
effect of land use on these wetlands (Hauenstein et al. 1988,
Ramírez et al., 1991).
The vectors used in the nMDS (Figure
9) as indicators of disturbance show a disturbance gradient from
the right-bottom corner (low disturbance) to the left-top corner (high
disturbance). According to this gradient, wetland number 3 has the lowest
disturbance level, while wetlands
number 10 and 7 have the highest level. At the high disturbance end
of the gradient species number becomes also high, together with the
importance of introduced species. These results contradict the findings
of Ramírez et al., (1992), who observed a reduction in the species
richness with disturbance. They also contradict the "intermediate disturbance
hypothesis" (Connell 1978) that predicts a unimodal response by the
species number (Grace 1999). But this interpretation does not represent
a total agreement with the reality: wetland number 8, for example, is
the only one with permanent water. On the other hand, wetland number
3 is intensively used as a watering place for cattle which implies a
high level of eutrophication. According to our observations in 1996
and 1997, wetlands 3 and 9 endured a longer flooding period than the
rest. Also, in wetland number 3 the aquatic plants reached their highest
importance. Because of this, it is probable that the above-mentioned
gradient in reality indicates a gradient of seasonality rather than
disturbance, starting in the right-bottom corner (near to permanent
flooding) moving to the left-top corner (near to terrestrial). This
suggestion is confirmed by the location of the plant communities in
the ordination diagram. Moreover, following Crow (1993), diversity increases
as a response to an alternating wet-dry climate, explaining the increase
in species number with seasonality in the ordination diagram.
The proportion of introduced species was directly correlated
with alpha diversity (R2 = 0.716, p < 0.01), however, its effects
on the beta diversity actually have an homogenization effect. The homogenization
index of Qian y Ricklefs (2006) was calculated, making all possible
pair-wise comparisons and calculating the Jaccard index of total species
composition (Jtotal) and the composition of native species (Jnative).
The homogenization index (H) results from the following formula: H =
Jtotal - Jnative. About 60 % of all these comparisons had negative values
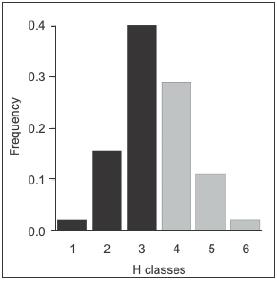
(Figure 10), which indicates that there was a weak
tendency to homogenization through introduced species.
|
| |
Figure
10. Distribution of homogenization indices (H) among pairs of
wetlands. Black bars represent differentiation effect, while grey
bars represent homogenization effect. H classes: 1 (-0.15 to -0.1),
2 (-0.1 to -0.05), 3 (-0.05 to 0), 4 (0 to 0.05), 5 (0.05 to 0.1)
and 6 (0.1 to 0.5).
Figura 10. Distribución de los índices de homogenización (H) comparando
pares de lagunas. Las barras negras representan efectos de diferenciación,
mientras que las barras grises representan efectos de homogenización.
Clases de H: 1 (-0.15 a -0.1), 2 (-0.1 a -0.05), 3 (-0.05 a 0),
4 (0 a 0.05), 5 (0.05 a 0.1) y 6 (0.1 a 0.5). |
CONCLUSIONS
It is concluded that the area of the studied seasonal
wetlands does not play any role in the determination of their species
richness or the number of communities present in them. The within wetland
diversity was also responsible for the high heterogeneity in the species
compositions of the wetlands.
The floristic similarity between wetlands (beta diversity)
was not related to the distance between them. This may be due to their
heterogeneity and the lack of restrictions for broad dispersal of the
plant species, at least at the geographical scale of this work.
It is difficult to estimate the effect of disturbance
on the species richness using ecological indicators. In this study,
the nMDS ordination did not reflect all aspects of disturbance in the
ordination diagram, relating to the importance of hemicryptophytes,
therophytes and introduced species. It is possible that factors not
measured in this research, like seasonality and the nutrient content
of soil and water, were also related to disturbance and important in
the determination of the species composition of the wetlands.
Finally, the importance of introduced plants was directly
correlated with the species richness of the wetlands; together they
produced a weak homogenization of the species composition between wetlands.
The high importance of introduced species and the low
amount of ephemeral wetlands specialists confirms their secondary, anthropogenic
character.
ACKNOWLEDGEMENTS
This work was supported by Grant TS3*CT94-03535 from
the E. C. and the Project F-95-06 from the research fund of the Universidad
Austral de Chile. The authors are indebted to and wish to thank D. Allworth
for help in the preparation of the manuscript. We also would like to
thank anonymous referees for their valuable comments.
REFERENCES
ALVAREZ, M. 2008. Diasporenbank und Ökologie der Vegetation
temporärer Gewässer im Cholchol-Gebiet (9. Region, Chile). J. Cramer,
Berlin. Diss. Bot. 407: 87 p.
AMIGO, J.; RAMÍREZ, C. 1998. A bioclimatic classification
of Chile: woodland communities in the temperate zone. Plant. Ecol. 136:
9-26.
ARRHENIUS, O. 1921. Species and area. J. Ecol. 9: 95-99
BARBOUR, M.G.; SOLOMESHCH, A.I.; WITHAM, C.W.; HOLLAND,
R.F.; MACDONALD, R.L.; CILLIERS, S.S.; MOLINA, J.A.; BUCK, J.J.; HILLMAN,
J.M. 2003. Vernal pool vegetation of California: variation within pools.
Madroño 50: 129-146.
BARBOUR, M.G.; SOLOMESHCH, A.I.; HOLLAND, R.F.; WITHAM,
C.W.; MACDONALD, R.L.; CILLIERS, S.S.; MOLINA, J.A.; BUCK, J.J.; HILLMAN,
J.M. 2005. Vernal pool vegetation of California: communities of long-inundated
deep habitats. Phytocoenologia 35: 177-200.
BERNINGER, O. 1929. Wald und offenes Land in Süd-Chile.
J. Engelhorns, Stuttgart. Geogr. Abh. 3: 130 p.
BESOAIN, M. 1985. Los suelos. In: Tosso, J. (ed.) Suelos
volcánicos de Chile. INIA, Santiago. pp: 1-107.
BLISS, S.A.; ZEDLER, P.H.; KEELEY, J.E.; ARROYO, M.T.K.
1998. A floristic survey of the temporary wetlands in the Mediterranean-climate
region of Chile. In: McComb, A.J.; Davis, J.A. (eds.). Wetlands for
the future. Gleneagles, Adelaide. pp: 219-228.
BRAY, J.R.; CURTIS, J.T. 1957. An ordination of the
upland forest communities of southern Wisconsin. Ecol. Monogr. 27: 325-349.
BRUMMITT, R.K.; POWELL, C.E. 1992. Authors of plant
names. Kew Royal Botanic Gardens. 732 p.
CHAMBERS, P.A.; LACOUL, P.; MURPHY, K.J.; THOMAZ, S.M.
2008. Global diversity of aquatic macrophytes in freshwater. Hydrobiologia
595: 9-26.
CONNELL, J.H. 1978. Diversity in tropical rain forests
and coral reefs. Science 199: 1302-1310.
CROW, G.E. 1993. Species diversity in aquatic angiosperms:
latitudinal patterns. Aquat. Bot. 44: 229-258.
DANSEREAU, P.; LEMS, K. 1957. The grading of dispersal
types in plant communities and their ecological significance. Institut
Botanique de l'Université de Montréal. 52 p.
DEIL, U. 2005. A review on habitats, plant traits and
vegetation of ephemeral wetlands - a global perspective. Phytocoenologia
35: 533-705.
DI CASTRI, F.; HAJEK, E.R. 1976. Bioclimatología de
Chile. Santiago, Universidad Católica de Chile. 129 p.
DIERSCHKE, H. 1994. Pflanzensoziologie. Stuttgart.
Eugen Ulmer, 683 p.
ELLENBERG, H., MUELLER-DOMBOIS, D. 1966. A key to Raunkiaer
plant life forms with revised subdivisions. Ber. Geobot. Inst. ETH,
Stift. Rübel 37: 56-73.
FEOLI, E., SCIMONE, M. 1984. Hierarchical diversity:
an application to broad-leaved woods of the Apennines. Giorn. Bot. Ital.
118: 1-15.
GRACE, J.B. 1999. The factors controlling species density
in herbaceous plant communities: an assessment. Perspect. Plant. Ecol.
2: 1-28.
GRILLAS, P.; GAUTHIER, P.; YAVERCOVSKI, N.; PERENNOU,
C. 2004. Mediterranean temporary pools. Station biologique de la Tour
du Valat, Le Sambuc, vol. 1, 119 p.
HAJEK, E.R.; DI CASTRI, F. 1975. Bioclimatografía de
Chile. Santiago, Universidad Católica de Chile. 107 p.
HAUENSTEIN, E.; RAMÍREZ, C.; LATSAGUE, M.; CONTRERAS,
D. 1988. Origen fitogeográfico y espectro biológico como medida del
grado de intervención antrópica en comunidades vegetales. Medio Ambiente
9: 140-142.
HAUENSTEIN, E.; GONZÁLEZ, M.; PEÑA-CORTÉS, F.; MUÑOZ-PEDREROS,
A. 2002. Clasificación y caracterización de la flora y vegetación de
los humedales de la costa de Toltén (IX Región, Chile). Gayana Bot.
59: 87-100.
HOLLAND, R.F.; JAIN, S.K. 1981. Insular biogeography
of vernal pools in the central valley of California. Am. Nat. 117: 24-37.
JACCARD, P. 1912. The distribution of the flora in
the Alpine zone. New Phytol. 11: 37-50.
KIRSCHNER, J. 2002. Species Plantarum (Juncaceae).
Australian Biological Resources Study, Canberra, vols. 7-8.
KRUSKAL, J.B. 1964. Nonmetric multidimensional scaling:
a numerical method. Psychometrika 29: 115-129.
LEYER, I.; WESCHE, K. 2007. Multivariate Statistik
in der Ökologie. Springer Berlin. 221 p.
LOMOLINO, M.V.; WEISER, M.D. 2001. Towards a more general
species-area relationship: diversity on all islands, great and small.
J. Biogeogr. 28: 431-445.
MACARTHUR, R.H.; WILSON, E.O. 1967. The theory of island
biogeography. Lamarks in Biology, Princeton. 203 p.
MARTICORENA, C.; QUEZADA, M. 1985. Catálogo de la flora
vascular de Chile. Gayana Bot. 42: 1-147.
MATTHEI, O. 1995. Manual de las malezas que crecen
en Chile. Santiago. Alfabeta 545 p.
OBERDORFER, E. 1960. Pfanzensoziologische Studien in
Chile. J. Cramer, Weinheim. Flora et Vegetatio Mundi 2: 208 p.
OTERO, L. 2006. La huella del fuego, historia de los
bosques nativos, poblamiento y cambios en el paisaje del sur de Chile.
Santiago. Pehuén 171 p.
POTT, R.; REMY, D. 2000. Gewässer des Binnenlandes.
Stuttgart, Eugen Ulmer. 255 p.
QIAN, H.; RICKLEFS, R.E. 2006. The role of exotic species
in homogeneizing the North American flora. Ecol. Lett. 9: 1293-1298.
RAMÍREZ, C. 1984. Einfluss der Jahreszeit auf Vegetationsaufnahmen
von Rasengesellschaften mit Thero- und Geophyten. In: Knapp, R. (ed.).
Sampling methods and taxon analysis in vegetation science. Hague, Dr.
W. Junk. pp: 181-183.
RAMÍREZ, C.; FERRIERE, F.; FIGUEROA, H. 1983. Estudio
fitosociológico de los bosques pantanosos templados del sur de Chile.
Rev. Chil. Hist. Nat. 56: 11-26.
RAMÍREZ, C.; ALVAREZ, M.; SAN MARTÍN, C. 2003. Diásporas
y mecanismos de dispersión en praderas antropogénicas de la X Región
de Los Lagos, Chile. Revista Geográfica de Valparaíso 34: 203-218.
RAMÍREZ, C.; SAN MARTÍN, C.; CONTRERAS, D. 1998. Diversidad
florística y vegetacional pratense en vegas, colinas y serranías al
poniente de Temuco, Chile. Ciencia e Investigación Agraria 25: 27-50.
RAMÍREZ, C.; SAN MARTÍN, C.; MAC DONALD, R. 1992. El
paisaje vegetal como indicador de cambios ambientales. Ambiente y Desarrollo
8: 67-71.
RAMÍREZ, C.; SAN MARTÍN, C.; OJEDA, P. 1997. Muestreo
y tabulación fitosociológica aplicados al estudio de los bosques nativos.
Bosque 18: 19-27.
RAMÍREZ, C.; FINOT, V.; SAN MARTÍN, C.; ELLIES, A.
1991. El valor indicador ecológico de las malezas del centro-sur de
Chile. Agro Sur 19: 94-116.
RAMÍREZ, C.; HAUENSTEIN, E.; CONTRERAS, D.; SAN MARTÍN,
J. 1988. Degradación de la vegetación en la depresión intermedia de
la Araucanía, Chile. Agro Sur 16: 1-14.
RAMÍREZ, C.; SAN MARTÍN, C.; CONTRERAS, D.; SAN MARTÍN,
J. 1994. Estudio fitosociológico de la vegetación pratense del valle
del río Chol-Chol (Cautín, Chile). Agro Sur 22: 41-56.
RAMÍREZ, C.; SAN MARTÍN, J.; HAUENSTEIN, E.; CONTRERAS,
D. 1989. Estudio fitosociológico de la vegetación de Rucamanque (Cautín,
Chile). Studia Botanica 8: 91-115.
R DEVELOPMENT CORE TEAM. 2005. R: a language and environment
for statistical computing. R Foundation for Statistical Computing, Viena.
http://www.R-project.org
RIVAS-MARTÍNEZ, S. 1993. Bases para una nueva clasificación
bioclimática de la tierra. Universidad Complutense de Madrid. Folia
Bot. Matritensis 10: 23 p.
SAN MARTÍN, J.; RAMÍREZ, C. 1986. Los bosques de ruil
de Chile central: sus especies vegetales y sus formas de vida. Maule
UC 10: 85-91.
SAN MARTÍN, C.; RAMÍREZ, C.; OJEDA, P. 1998. La vegetación
de lagunas primaverales en las cercanías de Temuco (Cautín, Chile).
Acta Bot. Malacitana 23: 99-120.
SCHMITHÜSEN, J. 1956. Die räumliche Ordnung der chilenischen
Vegetation. In: Troll, C.; Bartz, F.; Hahn, H. (eds.). Forschungen in
Chile. Geographisches Institut der Universität Bonn. pp: 1-86.
SCULTHORPE, C.D. 1967. The biology of aquatic vascular
plants. London, Edward Arnold. 610 p.
TUTIN, T.; BURGES, N.; CHATER, A.; EDMONSON, J.; HEYWOOD,
V.; MOORE, D.; VALENTINE, D.; WALTERS, S.; WEBB, D. 1996. Flora Europaea
UK, Cambridge University, 5 vols.
WHITTAKER, R.H. 1972. Evolution and measurement of
species diversity. Taxon 21: 213-251.
WOODLAND, D.W. 1991. Contemporary plant systematics.
New Jersey, Prentice Hall. 582 p.
ZEDLER, P.H. 1990. Life histories of vernal pool vascular
plants. In: Ikeda, D.H.; Schlising, R.A. (eds.). Vernal pool
plants - their habitat and biology. Herbarium CSU Chico, California,
pp: 123-146.
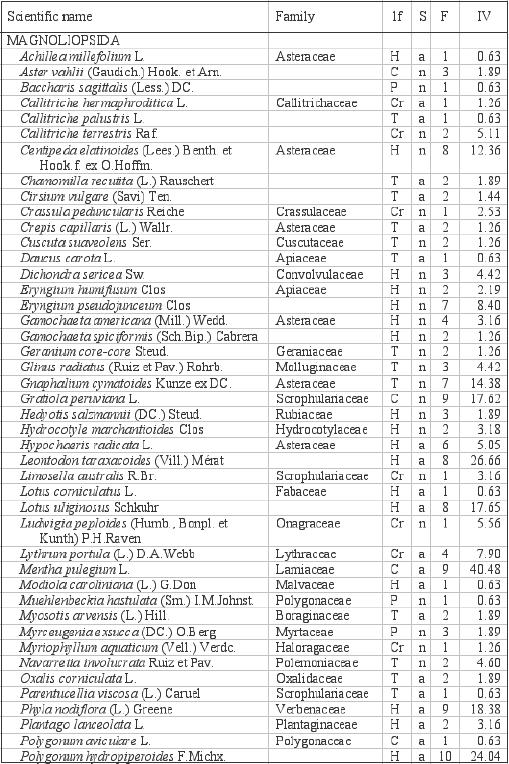
Appendix:
Scientific names, families, life-forms (lf), status (S), frequencies
in the pools (F), and importance values (IV) of the flora of the
anthropogenic vernal pools (Temuco, Chile). |
| |
 |
| |
| Appendix
(continuation) |
| |
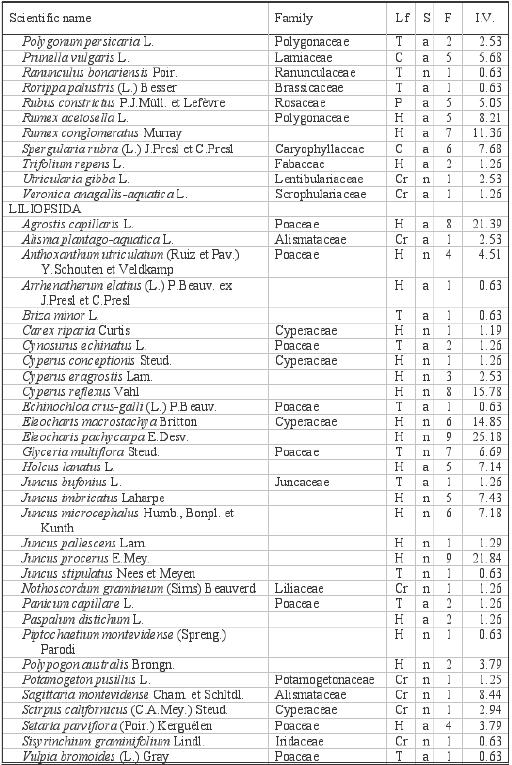 |
Life forms: phanerophytes (P), chamaephytes
(C), hemicryptophytes (H), cryptophytes (Cr) and therophytes
(T). Status: natives (n), introduced (a)
|