CIENCIAS ANIMALES: ANIMAL SCIENCE
Grazing behavior of two breeds of ewes on a secondary ecological succession of the Chiloé Island
Comportamiento en pastoreo de dos razas de ovejas sobre una sucesión ecológica secundaria de la Isla de Chiloé
Gallardo, M.A.a* Rivero, J.b, Triviños, M.I.c, Letelier, C.c
a Escuela
de Graduados, Facultad de Ciencias Veterinarias, Universidad Austral de Chile,
Valdivia, Chile.
b Instituto de Producción Animal, Facultad de
Ciencias Agrarias, Universidad Austral de Chile, Valdivia, Chile.
c Instituto de Ciencia Animal, Facultad de Ciencias Veterinarias,
Universidad Austral de Chile, Valdivia, Chile.
*Corresponding author:
María Asunción Gallardo E-mail address: mugallar@gmail.com
ABSTRACT
The aim of this experiment was to determine the grazing behavior of the Chilota and Suffolk Down breeds of sheep on a secondary ecological succession (such as a calafatal) in Chiloé. The experiment was performed between October and December 2011. Eight Chilota and 6 Suffolk Down ewes (intraracial similar weight and body condition) and their lambs, were selected and marked with an identification number on their backs. Three 1 ha paddocks located within a calafatal were used. Data recording was performed during 24 h, once a month, for 3 consecutive months by direct observation and manual recording of the following activities: grazing, standing, lying ruminating, standing ruminating, lying, walking and browsing. In this experiment, the Chilota ewes showed longer grazing times than the Suffolk Down ewes, despite their lower requirements and body size, spending less time walking at night than the Suffolk Down ewes. Also, the probability of occurrence of grazing behavior in this breed was higher than in Suffolk Down ewes. Monthly data indicated that the Chilota ewes showed shorter standing times (in October) and longer grazing times (in December) than the Suffolk Down ewes, according to a higher pasture availability. In summary, the Chilota ewes showed longer grazing times than the Suffolk Down ewes, spending less time standing ruminating during daylight or walking at night than the Suffolk Down ewes.
Keywords: Chilota, Suffolk Down, grazing, browsing
RESUMEN
El objetivo de este experimento fue determinar el comportamiento en pastoreo de las razas ovinas Chilota y Suffolk Down sobre una sucesión ecológica secundaria (como un calafatal) en la Isla de Chiloé. El experimento fue realizado entre octubre y diciembre de 2011. Ocho ovejas Chilota y seis ovejas Suffolk Down (de similar peso y condición corporal intraracial) y sus corderos, fueron seleccionados y marcados con un número de identificación en sus espaldas. Se utilizaron tres potreros de 1 ha ubicados dentro de un calafatal. El registro de los datos se realizó durante 24 h una vez al mes, por 3 meses consecutivos mediante observación directa y registro manual de las siguientes actividades: pastoreo, parada, rumiando echada, rumiando parada, echada, caminando y ramoneando. En este experimento las ovejas Chilota mostraron tiempos de pastoreo más largos que las ovejas Suffolk Down, a pesar de sus menores requerimientos y tamaño corporal, gastando menos tiempo caminando en la noche que las ovejas Suffolk Down. Además, la probabilidad de ocurrencia del comportamiento en pastoreo en esta raza fue mayor que en las ovejas Suffolk Down. Los datos mensuales indicaron que las ovejas Chilota mostraron menores tiempos de actividad parada (en octubre) y mayores tiempos de pastoreo (en diciembre) que las ovejas Suffolk Down, acorde a la mayor disponibilidad de pradera. En resumen, las ovejas Chilota mostraron tiempos de pastoreo más largos que las ovejas Suffolk Down, gastando menos tiempo rumiando durante las horas de luz o caminando en la noche que las ovejas Suffolk Down.
Palabras clave: Chilota, Suffolk Down, pastoreo, ramoneo.
INTRODUCTION
The Chiloé Island is part of the archipelago Chiloé and is located between 41° 44' and 43° 17' S, 72° 45' and 74° 30' W, Southern Chile. The Archipielago is composed of 40 islands that occupy a land area of 9,181.6 km2 (Montiel, 2003; Vera, 2003). The climate in Chiloé is maritime temperate, with rainfall ranging from 1,200 to 3,000 mm/year (average rainfall 2,070 mm/year). The annual relative humidity average ranges between 80 and 85%, with low frequency of frost. The mean annual temperature is 10.7 °C, (maximum of 30 °C in summer and minimum of 5 °C in winter) (Montiel, 2003). Soils are classified as Andisols, with low pH, high levels of exchangeable aluminum and low levels of phosphorus, nitrogen and potassium. The resultant of this ecological condition is a climax community that has been defined as an evergreen rainforest of Nothofagus dombeyi (De la Barra, 2008).
Sheep were introduced to the Archipelago in 1568, during the Spanish colonization. The introduced animal population was forced to survive under sub nutrition conditions, because of the poor nutritive quality of the existing vegetation and the hard climatic conditions (high humidity, constant rains, low temperatures and permanent winds) (De la Barra, 2008). The adaptation process to the agroecological conditions, along with the long period of isolation and inbreeding, led to a genetic and ethnological differentiation of the subsequent generations of animals, which are currently recognized as the Chilota breed of sheep (De la Barra, 2008; De la Barra et al., 2010).
The Chilota breed has low body weight, suggesting processes of adaptation to low temperature conditions and shortages of food. Also, Chilota's rusticity via resistance to Foot Rot and gastrointestinal parasites has been reported (Martínez et al., 2012).
The main feed resource for domestic and wild ruminants in the archipelago Chiloé is the calafatal, a secondary succession derivate from the anthropic intervention of the native forest, coupled with sheep introduction during the early stages after alteration, exposed to low grazing intensities (GI) (2 o. e.1 /ha/year). This system is dominated by shrubs such as Berberis darwinii and B. buxifolia, some ferns such as Blechnum penna marina and a herbaceous stratum, with species such as Agrostis capillaris, Holcus lanatus, Anthoxanthum odoratum, Trifolium dubium, Trifolium repens, Plantago lanceolata, Hypochaeris radicata, Lotus uliginosus, Lupinus sp., Madia sativa, Prunella minor and Carex sp. between others (De la Barra, 2008).
Regarding to the breed effect, Gallardo et al. (2014a) attempted to characterize the grazing behavior in ewes during daylight hours (09:00 to 20:00 h), reporting that the Chilota and the Suffolk Down ewes tended to graze mainly in the morning and evening hours. However, the browsing activity on this secondary succession dominated by shrubs, tended to be recorded more often in the Chilota than the Suffolk Down ewes. This behavior could be assigned to the greater rusticity of the Chilota sheep as the result of the adaptation process to the island conditions, implying that Chilota sheep can eat high fiber resources and could constitute an alternative to solve the problem of uncontrolled bush growth (due to inadequate GI) in the archipelago Chiloé.
The aim of this study was to identify different characteristics of the Chilota ewes (compared to Suffolk Down pattern) regarding to its grazing behavior when grazing a calafatal of Chiloé during 24 h.
MATERIAL AND METHODS
Study design
The experiment was conducted at the Butalcura Experimental Research Station of the Instituto de Investigaciones Agropecuarias (INIA-Chiloé), located in the Chiloé archipelago, during October, November and December 2011.
Animals
Fourteen healthy third lambing ewes, eight Chilota breed and six Suffolk Down ewes (and their lambs) live weight (LW) 43.77 ± 1.50 kg for the Chilota and 52.48 ± 2.54 kg for the Suffolk Down ewes and body condition score (BCS) 2.59 ± 0.04 and 2.75 ± 0.09 respectively for both breeds, were used. Considering the assay was performed immediately from the entrance of the animals to the paddocks and only for 24 h each, significant differences between breeds regarding LW or BCS were compensated by different number of animals per breed.
The ewes were randomly selected from a large free-grazing flock of Chilota and Suffolk Down sheep. All ewes were marked with an identification number on their backs. Three 1-ha paddocks within a calafatal were used. The behavior assay lasted 3 d, and previously, a 10 d adjustment period was necessary to get the ewes used to the experimental conditions.
Chemical composition of the pastures
At the beginning of each experimental period (October, November and December), a 1-ha experimental area of calafatal pasture was selected from five 1-ha paddocks. Three samples (consisting of 5 pooled sub-samples each) cut to soil level (0.10 m2 each) were taken from each 1-ha area in each experimental period.
The chemical analyses of the calafatal pasture were made in the laboratory of the Animal Production Institute of the Universidad Austral de Chile, to determine dry matter content (DM), crude protein (CP) (AOAC, 1996), metabolizable energy (ME), which was estimated by regression using a "D" value (Digestible organic matter/DM x 100) assessed in vitro (Tilley and Terry 1963), according to Goering and Van Soest (1970), neutral detergent fiber (NDF), according to Van Soest et al. (1991) and finally, acid detergent fiber (ADF), according to (AOAC, 1996).
Grazing behavior study
The ewes (separated by breed using mesh dividers), were located to graze a calafatal pasture divided into three paddocks (1.0 ha each), which were rotated every 5 days, because of low DM availabilities found in the calafatal (measured at ground level) during October, November and December 2011 (475, 584 and 763 kg ha-1, respectively).
Animals had free access to water in all the paddocks. Data collection was done by scan sampling every 10 minutes during daylight hours (from 09:00 to 21:00), and every 15 minutes at night (between 21:00 to 09:00 h). The evaluation was repeated once a month, for three consecutive months (October, November and December), performing an exhaustive recording of the following activities: grazing (ewe consuming forage from the pasture), standing (ewe standing, without eating or ruminating), lying-ruminating (ewe lying and ruminating), standing-ruminating (ewe standing and ruminating), lying (ewe lying), walking (ewe walking) and browsing (ewe consuming leaves and branches from the shrubs). Environmental temperature was recorded on the weather station existing at the Butalcura Experimental Research Center.
Statistical analyses
Results are supported by descriptive statistics to assess the most expressed behavior during observation hours. Additionally, to compare breeds, statistical inference of the time taken by a given behavior was done by repeated measures ANOVA, using SAS (2002-2003) statistical software. Simultaneously, data were analyzed using logistic regression (Hilbe, 2009) to estimate odds ratio and quantify the probability of occurrence of a given behavior. The chemical composition data were analyzed by analysis of variance, using SAS (20022003) statistical software and least square means were estimated and multiple range test were performed to quantify significant differences between breeds.
To quantify the breed effect the following statistical linear model was used:
Yij = Bi + eij
Where:
Yij = is the
amount of time that the jth ewe of the ith breed was observed
in a given behavior.
Bi = is the fixed effect of the ith breed.
eij = is the random residual
term ~N (0, σ2)
RESULTS
Ewes LW at the beginning (October), middle (November) and the end (December) of the experiment were 43.77 ± 1.50, 46.96 ± 1.88 and 47.37 ± 1.49 kg LW for the Chilota ewes, and 52.48 ± 2.54, 57.33 ± 2.28 and 58.25 ± 2.10 kg LW for the Suffolk Down ewes, respectively. Body condition score in October, November and December was 2.59 ± 0.04, 2.80 ± 0.09 and 3.34 ± 0.08 for the Chilota ewes and 2.75 ± 0.09, 2.71 ± 0.10 and 3.42 ± 0.19 for the Suffolk Down ewes, respectively.
Table 1 shows the chemical composition of the calafatal pasture2. December was the month with the lowest DM proportions, but higher CP, NDF (as much as October) and ME when compared with other months of the study.
Table 1.
Chemical composition of the calafatal during October, November and December
(a)
Cuadro 1. Composición química del calafatal durante octubre,
noviembre y diciembre (a)
(a) Different lower
case letters (a, b, c) denote significant effect of the month within the
pasture (P < 0.05)
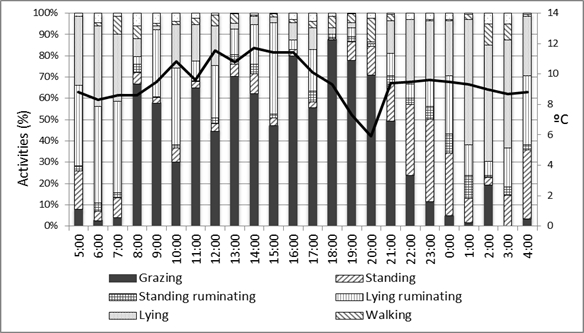
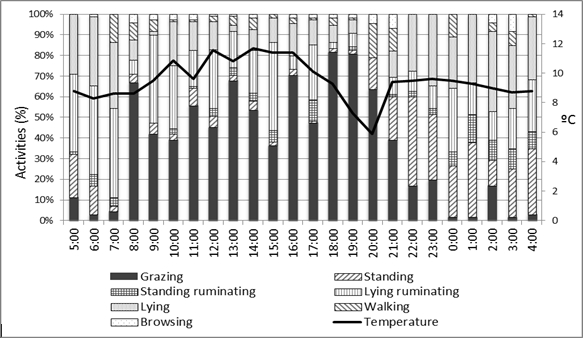
In relation to the grazing behavior study, during daylight hours (09:00 to 21:00 h), the main activity registered for the Chilota (Figure 1) and the Suffolk Down ewes (Figure 2) was grazing (until 21:00 h) and the second was lying-ruminating (until 17:00 h), when the environmental temperature was between 8 and 12 °C. From 18:00 to 20:00 h there was a decrease on the environmental temperature, which was coincident with a decrease in the grazing activity. The main activities registered at night (21:00 to 09:00) were lying and standing, when the environmental temperature was between 8 and 10 °C. Drinking activity was not recorded, because it was sporadic (Guzman, 2005; Olivares et al., 2009).
Figure 1. Main daily activities
of the Chilota ewes during October, November and December,
according the environmental temperature.
Figura 1. Principales actividades
diarias de las ovejas Chilota durante octubre, noviembre y
diciembre, acorde a la temperatura ambiental.
Figure 2. Main daily activities
of the Suffolk Down ewes during October, November and
December, according the environmental temperature.
Figura 2. Principales actividades
diarias de las ovejas Suffolk Down durante octubre,
noviembre y diciembre, acorde a la temperatura ambiental.
Table 2 shows the estimated Odds ratio (OR) and their 95% confidence intervals for the breed effect on the ewes (Summary Table). The probability of finding the Chilota ewes grazing was 1.14 (P < 0.01) times higher than the probability of finding the Suffolk Down ewes performing the same activity. However, the probability of lying ruminating activity was higher in the Suffolk Down than the Chilota ewes (P < 0.05).
Table 2.
Estimated odd ratios (OR) and their 95%
confidence intervals for the breed effect in ewes
(Summary Table)
Cuadro 2. Odd ratios estimados (OR)
y sus intervalos
de confianza al 95% para el efecto raza en las ovejas
(Tabla resumen)
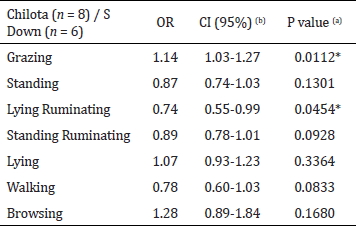
(a) Asterisk (*)
denote significant differences between
breeds
(b) CI: confidence interval
The grazing behavior of the Chilota and the Suffolk Down ewes, during 24 h, daylight hours (from 09:00 to 21:00), and at night (from 21:00 to 09:00) is shown in Table 3. During 24 h recording, the grazing times were longer (P < 0.05) in the Chilota than the Suffolk Down ewes. However, the walking times were longer (P < 0.05) in the Suffolk Down than the Chilota ewes. During daylight hours, the grazing activity times were longer in the Chilota than the Suffolk Down ewes. However, the standing ruminating times were longer for the Suffolk Down than the Chilota ewes. At night, walking activity times were longer in the Suffolk Down than the Chilota ewes.
Table 3.
Grazing behavior of the Chilota and the Suffolk Down ewes during 24 h, daylight
hours (from 09:00 to 21:00 h) and at night (from 21:00 to 09:00 h) (a)
(b)
Cuadro 3. Comportamiento en
pastoreo de las ovejas Chilota y Suffolk Down durante 24 h, las
horas luz (desde las 09:00 a las 21:00 h) y en la noche (desde las 21:00 a las
09:00 h) (a) (b)

(a) Results are the
mean of three consecutive measures (during October, November and
December); (b) Asterisk (*) denote significant effect of the breed
(P < 0.05)
Table 4 shows the grazing behavior of the Chilota and the Suffolk Down ewes during 24 h, in October, November and December. The Chilota ewes increased their grazing activity times from October to December, however the Suffolk Down ewes did not. The standing and lying activity times decreased from October to December in both breeds of ewes. In October, standing times were longer in the Suffolk Down than the Chilota ewes. In November, no significant differences between breeds regarding to behaviors were observed. Finally, in December, the grazing activity times were longer in the Chilota than the Suffolk Down ewes.
Table 4.
Grazing behavior of the Chilota and the Suffolk Down ewes, during October,
November and December (a)
Cuadro 4. Comportamiento en pastoreo de las ovejas Chilota y Suffolk Down,
durante
octubre, noviembre y diciembre (a)
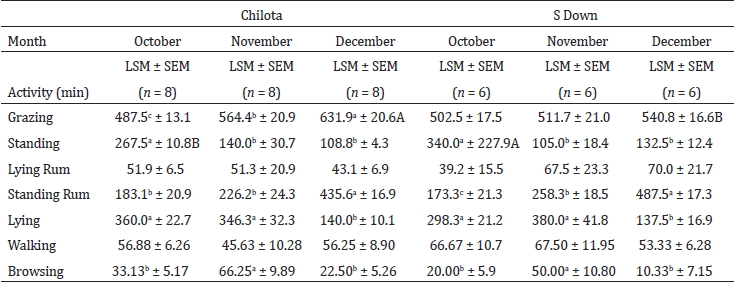
(a) Different small
letters (a, b, c) denote significant month effect (P < 0.05); capital
letters (A, B) denote significant effect of the lamb breed within the same month
(P < 0.05)
DISCUSSION
Many natural areas of the Chiloé Archipelago have been traditionally grazed by small cattle and sheep herds. Traditional grazing is considered a useful tool for keeping plant biodiversity in shrubland and wooded areas (Bartolomé et al., 2000; Papachristou et al., 2005). Only an unbiased use of all species in a community could maintain the plant biodiversity under high herbivore pressure (Provenza et al., 2003; Dziba et al., 2007). Thus, sheep being very social animals may modify their foraging behavior, influencing not only the place where animals feed, but also causing an impact on food resource (Zanine et al., 2006; Sibbald et al., 2008).
In this study, a trend to graze between 08:00 and 21:00 h was observed, independent of the environmental temperature, however, a progressive decrease in the grazing activity from 19:00 h was registered, coincident with a gradual decrease of the environmental temperature from 16:00 h.
Irrespective of the month, animals avoided grazing in darkness (Hessle et al., 2008; Gallardo et al., 2014b). Lin et al. (2011), evaluating the effect of different GI on the grazing behavior in a tail fat breed of sheep, reported the existence of two mayor grazing periods, a shorter in the morning and a longer grazing period in the afternoon (Fierro and Bryant, 1990; Wang, 1997). This could be explained by a greater preference of the ruminants to consume food in the afternoon (Fisher et al., 1999, 2002), due to its greater digestibility compared with the fodder harvested in the morning (Burns et al., 2005; Huntington and Burns, 2008). Also, Olivares et al. (2009) reported that the sheep presented a uniform distribution of the activities during daylight hours, with two peaks of grazing, one in the morning and another in the afternoon.
In this study, a synchronous behavior within each monthly measurement was observed, which increased as the months passed and the animals became used to the experiment, and the quality and pasture availability increased.
Independent of the measuring month, at night time hours, both breeds of ewes were standing, standing ruminating or simply lying, for shelter, avoiding grazing in the dark due to predation risk (Hessle et al., 2008) and also, as a form of energy saving. These results agree with those obtained by Gautrais et al. (2007), who reported that under domestication conditions, the animals tend to repeat behaviors, being more active at the time of great changes, as dawn and dusk and least active at midday or midnight. De Moura Zanine et al. (2006), working on pastures with different morphological structure, reported that the sheep and the goats grazed between 06:00 and 11:00 h with peaks of grazing and rumination in hours of mild temperatures, remaining at resting in hours of very high temperatures (Guzmán, 2005; Zanine et al., 2006; Olivares et al., 2009).
Regarding the grazing times during 24 h, they were longer in the Chilota breed than the Suffolk Down ewes, as a consequence of the significant differences in the grazing times between both breeds of ewes during daylight hours. Also, the greater walking behavior times recorded for the Suffolk Down ewes during 24 h, could be a consequence of their greater walking behavior times at night. Olivares et al. (2009), comparing the breed effect in sheep in three phenological stages (vegetative, reproductive and dry), reported that the main activity recorded was foraging for all stages, a finding consistent with those reported by several authors (Caris, 2004; Animut et al., 2005; Guzmán, 2005; Schlecht et al., 2006). Also, Olivares et al. (2009), working with Merino and Suffolk Down sheep and Boer goats in a wheatgrass pasture, assessed the grazing behavior for three consecutive days in three phenological stages of grassland, concluding that the main activity recorded was foraging, without finding any significant differences between sheep, but between sheep and goats.
Contrary to Fierro and Bryant (1990), the grazing activity times were positively associated with the ME contents. This behavior was observed in both breeds, with a trend to increase the forage intake when pasture ME content increased. Thus, in December (month which the calafatal showed the highest ME content) and as a result of a longer grazing activity times during daylight hours, instead of their lower size and requirements (Webster, 1985; De la Barra, 2008) the Chilota ewes spent more time grazing at 24 h and during daylight than Suffolk Down ewes. Also, the probability of finding Chilota ewes breed grazing was higher than Suffolk Down ewes, however, no significant differences between breeds were found in relation to the probability of walking behavior occurrence.
During daylight hours, the standing ruminating activity time was longer in the Suffolk Down than the Chilota ewes, but this behavior was not a consequence of a previous longer grazing or browsing times in the Suffolk Down ewes.
In this study, no significant differences on the browsing activity times between breeds, such as those reported by Gallardo et al. (2014b) in lambs grazing on a calafatal, were found.
After grazing activity, the presence of an arboreal and shrub layer, could be a decisive factor on lying (in both breeds) or lying ruminating (higher probability for the Suffolk Down than the Chilota ewes), protecting them from the intense summer sun at high temperatures hours, because this behavior also varies in response to environmental changes. Also, Caris (2004), observing the sheep with or without presence of shrubs (Acacia caven), reported that for these animals the most important activity within the day was eating, and the presence of a tree layer as a protection had an influence on drinking, resting and walking activities. Others factors such as day type (sunny and cloudy) or the physiological condition of the animals, also determined the amount of time spent on each activity, being eating the most important activity reported (Penning et al., 2004).
These results show that the grazing behavior appears to be more affected by the season than the breed of ewe (Hessle et al., 2008), probably due to the increased quality and pasture availability. Lin et al. (2011) reported that the sheep increased or maintained their grazing behavior times by decreasing resting times, to avoid the effect of shorting daylight duration (in the present experiment the same effect was observed as daylight hours and pasture supply increased from October to December). Also, values reported for the grazing activity during daylight hours were higher than those reported by Lin et al. (2011) working with a fat tailed breed of sheep during a daylight period (16 h), with GI 4 (moderate) and lower than those obtained with GI 5 (heavy).
Regarding the stocking rate (SR), Animut et al. (2005) studied its effect on the grazing behavior and energy expenditure (EE) in growing sheep and goat co-grazing grass/forb pastures, and reported that the effect of SR on the grazing behavior was similar in sheep and goat, but the effects of SR on EE can affect the average daily gain in both species. Also, Lin et al. (2011), reported that sheep exposed to high GI are able to increase their grazing and walking times, expending a lot of energy, which otherwise could be used for growth or production (Schutz and Jensen, 2001; Schutz et al., 2001). In the present experiment, SR was adequate to the conditions of the study area (calafatal of the Chiloé archipelago).
CONCLUSION
Under the conditions of this experiment, the Chilota ewes performed longer grazing activity times than the Suffolk Down ewes, despite their smaller body size and lower requirements, spending less time standing ruminating during daylight hours or walking at night than the Suffolk Down ewes.
ACKNOWLEDGEMENTS
To Tamara Tadich for her collaboration in this publication. Study financed by Graduated School, Faculty of Veterinary Science, Universidad Austral de Chile (UACh).
NOTAS
1 The ovine equivalent (o. e.) is a measure of the stocking rate corresponding to the energetic requirements of a sheep (55 kg LW) rearing a 100 day-old suckling lamb.
2 The botanical composition was included in Gallardo et al. (2014b).
REFERENCES
ANIMUT, G., GOETSCH, A.L., AIKEN, G.E., PUCHALA, R., DETWEILER, G., KREHBIE, C.R., MERKEL, R.C., SAHLU, T., DAWSOND, L.J. 2005. Grazing behavior and energy expenditure by sheep and goats co-grazing grass/forb pastures at three stocking rates. Small Ruminant Research 59: 191-201.
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS, 1996. Official Methods of Analysis, 16th ed. AOAC, Gaithersburg, VA, USA.
BARTOLOMÉ, J., FRANCH, J., PLAIXATS, J., SELIGMAN, N.G. 2000. Grazing alone is not enough to maintain landscape diversity in the Montseny Biosphere Reserve. Agriculture, Ecosystems and Environment 77: 267-273.
BURNS, J.C., MAYLAND, H.F., FISHER, D.S. 2005. Dry matter intake and digestion of alfalfa harvested at sunset and sunrise. Journal of Animal Science 83: 262-270.
CARIS Y. 2004. Conducta de ovinos en pradera anual de clima mediterráneo con y sin protección de espinos, Región Metropolitana. Memoria de título, Facultad de Ciencias Agrarias, Universidad de Chile. 55 p.
DE LA BARRA, R. 2008. Efecto de la introducción de la ganadería en el archipiélago de Chiloé, Chile. Tesis Doctoral, Departamento de Ecología y departamento de Producción Animal, Universidad de León. 202 p.
DE LA BARRA, R., URIBE, H., LATORRE, E., SAN PRIMITIVO, F., ARRANZ, J. 2010. Genetic structure and diversity of four Chilean sheep breeds. Chilean Journal of Agricultural Research 70: 646-651.
DE MOURA ZANINE, A., SANTOS, E.M., FERREIRA, D., LORA, A., LORA, G. 2006. Comportamento ingestivo de ovinos e caprinos em pastagens de diferentes estructuras morfológicas (intake behaviour of sheep and goat in pastures). Revista Electrónica de Veterinaria REDVET, 7(3). ISSN 1695-7504. <http://www.veterinaria.org/revistas/redvet/n030306/030610.pdf) (accessed Febraury 2013).
DZIBA, L., PROVENZA, F., VILLALBA, J., ATWOOD, S. 2007. Supplemental energy and proteinincrease use of sagebrush by sheep. Small Ruminant Research 69: 203-207.
FIERRO, L.C., BRYANT, F.C. 1990. Grazing activities and bioenergetics of sheep on native range in Southern Peru. Small Ruminant Research 3: 135-146.
FISHER, D.S., MAYLAND, H.F., BURNS, J.C. 1999. Variation in ruminants' preference for tall fescue hays cut either at sundown or at sunup. Journal of Animal Science 77: 762-768.
FISHER, D.S., MAYLAND, H.F., BURNS, J.C. 2002. Variation in ruminant preference for alfalfa hays cut at sunup and sundown. Crop Science 42: 231-237.
GALLARDO, M., PULIDO, R., DE LA BARRA, R., MARTINEZ M.E. 2014a. Pilot study for preferences and grazing behavior in two breeds of sheep on a secondary succession of the Chiloé Archipelago. Ciencia e Investigación Agraria 41 (2): 249-254.
GALLARDO, M.A., RIVERO, M.J., FAÚNDEZ, L. 2014b. The grazing behavior and diet selectivity of two lamb breeds on secondary successional pastures in the Chiloé Archipelago. Livestock Science 161: 69-79.
GAUTRAIS, J., MICHELENA, P., SIBBALD, A., BON, R., DENEU-BOURG, J.L. 2007. Allelomimetic synchronization in Merino sheep. Animal Behaviour 74: 1443-1454.
GOERING, H.K., VAN SOEST, P.J. 1970. Forage fiber analysis. Agriculture Handbook N° 379. ARS-USDA, Washington.
GUZMÁN, C. 2005. Influencia del estrato arbóreo (Acacia caven) en la conducta ambiental de ovinos en pastoreo, en el secano interior de la Región Metropolitana. Memoria de título. Facultad de Ciencias Agronómicas, Universidad de Chile. 56 p.
HESSLE, A., RUTTER, M., WALLIN, K. 2008. Effect of breed, season and pasture moisture gradient on foraging behaviour in cattle on semi-natural grasslands. Applied Animal Behaviour Science 111: 108-119.
HILBE, J.M. 2009. Logistic Regression Models. Chapman & Hall/CRC Press. ISBN 978-1-4200-7575-5.
HUNTINGTON, G.B., BURNS, J.C. 2008. The interaction of harvesting time of day of switchgrass hay and ruminal degradability of supplemental protein offered to beef steers. Journal of Animal Science 86: 159-166.
LIN, L., DICKHOEFER, U., MÜLLER, K., WURINA, SUSENBETH, A. 2011. Grazing behavior of sheep at different stocking rates in the Inner Mongolian steppe, China. Applied Animal Behaviour Science 129: 36-42.
MARTÍNEZ, M.E., CALDERÓN, C., URIBE, H., DE LA BARRA, R. 2012. Effect of management practices in the productive performance of three sheep breeds in the Chiloé Archipelago, Chile. Journal of Livestock Science 3: 57-66.
MONTIEL, D. 2003. Chiloé: Crónicas de un mundo insular. DIMAR, Puerto Montt.
OLIVARES, A., CASTELLARO, G., URRA, H., HIDALGO, J. 2009. Conducta ambiental de rumiantes menores en un pastizal de agropiro (Thinopyrum ponticum (Podp)). XXXIV Reunión Anual de la Sociedad Chilena de Producción Animal A. G. (SOCHIPA). Universidad Católica de Termico, 21-23 de Octubre de 2009, Pucón, Chile, pp. 183-184.
PAPACHRISTOU, T.G., PLATIS, P.D., NASTIS, A.S. 2005. Foraging behavior of cattle and goats in oak forest stands of varying coppicing age in Northern Greece. Small Ruminant Research 59: 181-189.
PENNING P.D., RUTTER S.M. 2004. Ingestive Behaviour. In: Penning, P.D. (Ed.), Herbage Intake Handbook, 2nd Ed. The British Grassland Society, Department of Agriculture, University of Reading, Reading, pp. 151-176.
PROVENZA, F.D., VILLALBA, J.J., DZIBA, L.E., ATWOOD, S.B., BANNER, R.E. 2003. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Ruminant Research 49: 257-274.
SAS (r) (Statistical Analysis System) 9.1. Copyright (c) 20022003 by SAS Institute INC., Cary, North Carolina.
SCHLECHT, E., HIERNAUX, P., KADAOURE, I., HULSEBUSCH, C., MAHLER, F. 2006. A spatio-temporal analysis of forage availability and grazing and excretion behavior of herded and free grazing cattle, sheep and goats in Western Niger. Agriculture, Ecosystems and Environment 113: 226-242.
SCHUTZ, K.E., JENSEN, P. 2001. Effects of resource allocation on behavioral strategies: a comparison of red jungle fowl (Gallusgallus) and two domesticated breeds of poultry. Ethology 107(8): 753-765.
SCHUTZ, K., FORKMAN, B.E., JENSEN, P. 2001. Domestication effects on foraging strategy, social behavior and different fear responses: a comparison between the red jungle fowl (Gallus gallus) and a modern layer strain. Applied Animal Behaviour Science 74(1): 1-14.
SIBBALD, A.M., OOM, S.P., HOOPER, R.J., ANDERSON, R.M. 2008. Effects of social behavior on the spatial distribution of sheep grazing a complex vegetation mosaic. Applied Animal Behaviour Science 115: 149-159.
TILLEY, J., TERRY, R. 1963. A two stages technique for the in vitro digestion of forages crops. Journal of the British Grassland Society 18: 104-111.
VAN SOEST, P.J., ROBERTSON, J., LEWIS, B. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal of Dairy Science 74: 3583-3595.
VERA, W. 2003. La agricultura Chilota: De una lenta transformación a una profunda transformación. Agraria Conseil General Finistere. Puerto Montt.
WANG, S.P. 1997. Behavior ecology of grazing sheep: II Influence of stocking rates on foraging behavior of whether. Acta Pratacultura Sinica 6: 10-17.
WEBSTER, A.J.F. 1985. Differences in the energetic efficiency of animal growth. Journal of Animal Science 61: 92-103.
ZANINE, A.M., SANTOS, E.M., FERREIRA, D.J., LORA, A., LORA, G. 2006. Comportamento ingestivo de ovinos e caprinos em pastagens de diferentes estruturas morfológicas. Revista Electrónica de Veterinaria (REDVET) 7(3): 1-10.
Received 12.09.14
Accepted 25.11.14