1 Facultad de Medicina Veterinaria, Universidad de Concepción, Casilla 537, Chillán, Chile. 2Facultad de Veterinaria, Universidad de Córdoba, España.
Biopsies of the Gluteus medius
muscle were taken at 3, 6, and 9 cm of depth in 16 inactive male and female
draught horses of the equine support plan, with ages ranging between 5 and 15
years old. The biopsies were analized by the histochemical techniques myosin
ATPasa, to determine the fibrilar composition, area and least diameter, and
the NADH-TR technique to determine the oxidative capacity of the fibres. Also,
the activity of the citrate sintetase (CS), lactate deshidrogenase (LDH) and
creatine phosphokinase (CK) enzymes were determined.
The average values in relation with the percentage of each fibre type were 33%,
28%, 17% and 22% for the fibres type I, IIA, IIB oxidative and IIB nonoxidative,
respectively. The average area of the fibres was 3959, 4823 and 7087 µm2
and the average minimum diameter was 47, 52 and 59 µm for fibres type
I, IIA, IIB respectively. The average of the activity of the enzymes CK, LDH
and CS was 0.484, 0.337 and 671 µmol/min/mg protein respectively, showing
an increase of the activity of CS with the depth of the biopsy. This increase
is related with the percentage of fibres type I. Results obtained show that
Chilean mixed draught horses have a high aerobic capacity and relative low anaerobic
capacity in the Gluteus medius muscle. These muscle characteristics are
consistent with the sort of work carried out by these animals: prolonged and
sustained exercise of low intensity.
Palabras claves: histoquímica, bioquímica, músculo, caballo.
Key words: histochemistry, biochemistry, muscle, horse.
INTRODUCCION
La mayoría de los músculos esqueléticos están compuestos por fibras con diferentes propiedades contráctiles y metabólicas, las cuales satisfacen requerimientos específicos de ellos. El uso de las técnicas histoquímicas ha permitido caracterizar estas fibras, siendo la clasificacion más usada para el caballo la que se basa en la actividad miosina ATPasa de la fibra muscular, tras preincubación en diferentes pHs. Según la actividad miosina ATPasa, es posible identificar fibras tipo I (contracción lenta), fibras tipo IIA y IIB (contracción rápida) (Brooke y Kaiser, 1970) y fibras tipo IIC que se presentan en músculos fetales y músculos de animales jóvenes (Essén-Gustavsson y col., 1980).
La técnica de biopsia muscular descrita por Lindholm y Piehl (1974) ha permitido estudiar la composición de la fibra muscular y su perfil bioquímico en el músculo del caballo y otras especies (Snow y Guy, 1976; Essén-Gustavsson y col., 1980). La mayoría de los estudios se han realizado en el músculo Gluteus medius debido a su gran volumen, fácil acceso y ausencia de estructuras vulnerables (López-Rivero y col., 1993). Sin embargo, también se han realizado estudios en los músculos Biceps femoris, Semitendinosus y Vastus lateralis (Guy y Snow, 1977; Lexell y col., 1983). Además, en las fibras se puede determinar su capacidad oxidativa utilizando la enzima NADH tetrazolium reductasa, encontrándose que las fibras tipo I y IIA tienen una elevada capacidad oxidativa; en cambio, en las fibras tipo IIB esta propiedad es variable (Essén-Gustavsson y col., 1980; López-Rivero y col., 1990).
La composición fibrilar de un músculo está determinada genéticamente y varía por factores tales como : la raza, el sexo, la edad y el entrenamiento a que es sometido el animal (Ronéus y col., 1991). Así, morfológicamente las fibras tipo I o "fibras rojas" son de menor tamaño que las fibras de tipo IIA y éstas a su vez son más pequeñas que las fibras tipo IIB (Essén-Gustavsson y col., 1989; López-Rivero y col., 1990). Teóricamente las fibras tipo I están mejor adaptadas para el metabolismo aeróbico (Essén-Gustavsson y col., 1984), por lo tanto, están bien dotadas para resistir la fatiga, por lo que resultan esenciales en actividades musculares de larga duración y baja intensidad. Por otra parte, las fibras tipo IIA, además de experimentar contracción rápida, tienen una considerable capacidad de resistencia, lo que las hace imprescindibles para caballos que corren grandes distancias (López-Rivero, 1995).
Las fibras tipo IIB "pálidas" producen alta cantidad de energía a través de vías anaeróbicas por tiempos cortos, siendo la acumulación de lactato más rápida en las fibras tipo IIB no oxidativas, lo que produce acidosis muscular y limita la capacidad para resistir la fatiga (Barlow y col., 1984).
Las fibras tipo IIC corresponderían a células indiferenciadas en estados de desarrollo, siendo su capacidad oxidativa y su tamaño intermedio entre las fibras tipo I y tipo II (Essén-Gustavsson y col., 1980).
Diversos estudios demuestran que el tipo de fibra varía dentro del músculo según la profundidad a que se tome la biopsia. Así por ejemplo, el porcentaje de fibras tipo I aumenta progresivamente con la profundidad del muestreo (Kline y col., 1987; López-Rivero y col., 1992). Observándose en las biopsias superficiales (2-3 cm de profundidad) un mayor porcentaje de fibras tipo IIB y en las más profundas (6 y 9 cm) un mayor porcentaje de fibras tipo I (Guy y Snow, 1980; Lexell y col., 1983; López-Rivero y col., 1992; Islas y col., 1996).
Respecto a los cambios bioquímicos que ocurren en la fibra muscular está demostrado que existen variaciones en la actividad de enzimas oxidativas y del metabolismo glicolítico según el tipo de fibra (Barlow y col., 1984; Essén-Gustavsson y Lindholm, 1985), siendo importante determinar entre otras las enzimas creatinafosfoquinasa (CK), deshidrogenasa láctica (LDH) y citrato sintetasa (CS), que participan en el metabolismo energético de la fibra muscular. La enzima citrato sintetasa representa el potencial oxidativo de la fibra muscular, aumenta con la edad y el entrenamiento en el caballo, lo que puede estar relacionado con el incremento de las fibras tipo I y IIA y un descenso de las fibras tipo lIB (Guy y Snow, 1977; Essén-Gustavsson y Lindholm, 1985; Hodgson y col., 1986; Ronéus y col., 1992).
La CK se encuentra presente en diversos tejidos; sin embargo, su mayor concentración se encuentra en el tejido muscular (Galitzer y Oehme, 1985) y junto con la LDH son enzimas evaluadoras del metabolismo glicolítico de la fibra muscular (Essén-Gustavsson y Lindholm, 1985).
Debido a que las características histoquímicas y bioquímicas de los músculos locomotores varían significativamente entre los equinos dependiendo del trabajo que ellos realizan, se postula que el equino mestizo de tiro presenta características musculares propias relacionadas con su origen y el tipo específico de trabajo que ellos realizan.
Esta investigación se diseñó con el propósito de determinar la composición fibrilar del músculo Gluteus medius, capacidad oxidativa, área, diámetro menor, y la actividad específica de algunas enzimas del metabolismo aeróbico y anaeróbico del músculo en equinos mestizos de tiro.
MATERIAL Y
METODOS
Se utilizaron 14 hembras y 2 machos equinos mestizos de tiro F1, clínicamente
sanos, producto de la cruza de hembras de tiro criollo con potros Bretones y
Ardanés del Plan de Fomento Equino, pertenecientes al fundo Pupunahue
de la X Región. Todos los animales no realizaron faenas agrícolas
durante 90 días previo a la obtención de las biopsias y recibieron
la misma alimentación en base a praderas naturales mejoradas y suplementados
con heno de trébol durante el invierno.
El peso promedio fue de 650 ± 40 kg y la edad fluctuó entre 5 y 15 años.
Biopsias musculares. Se obtuvieron dos muestras de biopsia del lado izquierdo del músculo Gluteus medius mediante aguja de biopsia de Lindholm y Piehl (1974) a 15 cm de la tuberosidad coxal en dirección dorsocaudal a la base de la cola. Se obtuvieron las muestras a 3, 6 y 9 cm de profundidad en el mismo sitio. Un trozo de tejido muscular se cubrió con OCT (Tissue tek, Miles Sci USA) y seguidamente se sumergió en isopentano en frío (-60° C) por 2-3 minutos. Posteriormente, las muestras se mantuvieron en nitrógeno líquido hasta su procesamiento. Otro trozo de tejido se guardó en tubos en nitrógeno líquido hasta la determinación de las enzimas.
Técnicas histoquímicas. Se obtuvieron 24 cortes de biopsias musculares de cada profundidad (3, 6 y 9 cm), de 10 µm de grosor en un crióstato (Micrón HM 500) a -20°C. Los cortes para determinar la actividad miosina ATPasa y actividad oxidativa fueron procesados según las técnicas descritas por Brooke y Kaiser (1970). Realizadas las técnicas, se tomaron dos microfotografías para cada profundidad de biopsia y para cada reacción enzimática, utilizando un fotomicroscopio marca Leitz con aumento 100x.
En las fotografías, realizadas en blanco y negro, se contó y se clasificó un total de 200 fibras musculares y se hizo el cálculo porcentual promedio de cada tipo de fibra con la técnica ATPasa. Posteriormente se calculó el área y diámetro menor de ellas por medio de un programa para análisis de imágenes por computación de la Facultad de Veterinaria de la Universidad de Córdoba, España. Los porcentajes de fibras oxidativas y no oxidativas se determinaron contando 100 fibras por biopsia teñida con la tinción nicotín amida tetrazolium reductasa (NADH-TR) (Brooke y Kaiser, 1970).
Determinaciones de actividades enzimáticas. Las biopsias fueron descongeladas sólo en el momento de procesarlas. En 50 mg de tejido muscular, previa extracción del tejido conectivo y graso, fue centrifugado 3 veces a 1.000 g en 5 ml de tampón fosfato pH 7.3 (PBS) frío. Posteriormente, el sedimento se solubilizó en 10 ml de PBS y se homogeneizó a 8.000 rpm en triturador Turex-X durante 30 segundos. Luego se sonicó por 20 segundos y se centrifugó a 7.000 g durante una hora a 4°C (Rudolph y col., 1993).
En el sobrenadante se determinaron las enzimas CK, LDH y CS. La actividad de
las enzimas CK (E.C. 2.3.2.2.) y LDH (E.C. 1.1.1.27) se realizó usando
métodos cinéticos con reactivos comerciales (Boehringer Mannheim)
y la actividad de la CS (E.C. 4.1.3.7.) se determinó según la
técnica descrita por Alp y col. (1976).
La determinación de las proteínas totales se realizó utilizando el método de Lowry y col. (1951).
Análisis estadístico. Los resultados fueron analizados estadísticamente, determinándose la media (X) y desviación estándar (DE) de cada variable y se realizó un análisis de varianza para determinar si las diferencias por efecto de la profundidad de la biopsia eran significativas. Además, se correlacionó el tipo de fibra con la actividad enzimática.
RESULTADOS
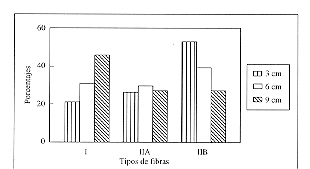
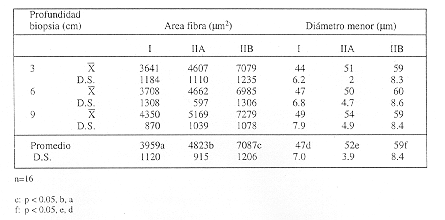
La distribución del porcentaje de fibras del músculo Gluteus medius en el equino mestizo de tiro mejorado adulto se presenta en la tabla 1. Se puede observar que el porcentaje promedio de fibras tipo I fue de 33% y aumenta con la profundidad de la biopsia de un 21% a 3 cm, 31% a 6 cm y 46% a 9 cm, siendo esta diferencia estadísticamente significativa (p<0.05) (fig. 1).
El porcentaje promedio de fibras tipo IIA fue de 28%, no presentando variaciones significativas al relacionarlas con la profundidad de la biopsia. El porcentaje promedio de fibras tipo IIB fue de un 40%, siendo de un 52% a los 3 cm de profundidad y disminuyendo a un 27% a 9 cm de profundidad, siendo esta diferencia estadísticamente significativa (p<0.05).
Al analizar la capacidad oxidativa de las biopsias musculares mediante la técnica de NADH-TR se determinó que el porcentaje promedio de fibras tipo IIB oxidativas fue de 17%, no existiendo diferencias a las distintas profundidades de muestreo (p>0.05) (tabla 1, fig. 2).
El porcentaje promedio de fibras tipo IIB no oxidativas fue de 22%, existiendo diferencias significativas entre la profundidad a 3 cm, respecto a la de 6 cm y a la de 9 cm (p<0.05) (tabla 1).
El área promedio de las fibras tipo I, IIA y IIB fue de 3.966, 4.830 y 7.052 µm2 respectivamente, no presentando variaciones significativas por efecto de la profundidad de la biopsia; sin embargo, las fibras tipo IIB presentan un área mayor (p>0.01) que las fibras tipo I y IIA en las tres profundidades de muestreo y las fibras tipo IIA a la profundidad de 9 cm presentan una mayor área que las fibras tipo I (p<0.05) (tabla 2).
Fibre types (%) of Gluteus medius muscle in inactive mixed draught horses presented as mean values and standard deviation.

inactive mixed draught horses at 3, 6 and 9 cm of depth of biopsy.
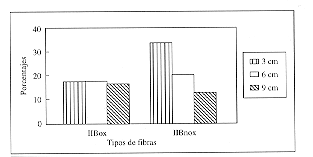
as mean values and standard deviation.
El diámetro menor promedio de las fibras tipo I, IIA y IIB fue de 47, 52 y 59 µm respectivamente y no presentó variaciones significativas por efecto de la profundidad de la biopsia, aunque las fibras tipo IIB tienen el diámetro menor más grande (p<0.05).
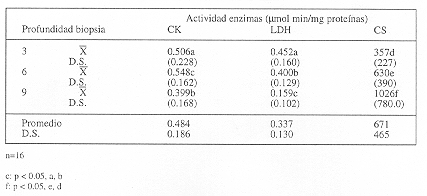
En relación a las actividades enzimáticas, se observó que la actividad promedio de las enzimas CK, LDH y CS fue de 0.484, 0.337 y 671 µmol/min/mg proteína, respectivamente (tabla 3).
El promedio de la actividad de CK a los 3 cm de profundidad fue de 0.506 µmol/min/mg proteína, la cual disminuyó a 0.399 µmol/min/mg proteína a los 9 cm de profundidad, siendo este descenso estadís-ticamente significativo (p<0.05) (tabla 3).
El promedio de la actividad de la LDH fue de 0.452 µmol/min/mg proteína, disminuyendo a 0.400 µmol/min/mg proteína a los 6 cm de profundidad y al 0.159 a los 9 cm de profundidad, siendo este descenso significativo respecto a los 3 y 9 cm de profundidad (p<0.05) (tabla 3).
El promedio determinado para CS a los 3 cm de profundidad fue de 357 µmol/min/mg proteína, la cual aumentó a 630 y 1.206 µmol/min/mg proteína a los 6 y 9 cm de profundidad, respectivamente, siendo este aumento significativo (p<0.05) (tabla 3).
El análisis de correlación entre el tipo de fibra y la actividad enzimática demostró que existe correlación positiva (p>0.05) solamente entre la actividad de la enzima CS y las tres profundidades de muestreo.
DISCUSION
El caballo mestizo de tiro es producto de la cruza de hembras de tiro criollo con potros Ardanés y Bretones finos pertenecientes al Plan de Fomento Equino. La composición fibrilar del músculo Gluteus medius en el caballo mestizo de tiro en condiciones de reposo no ha sido analizada previamente y los resultados se comparan con los obtenidos en el mestizo de tiro criollo y otras razas.
Los tipos de fibras encontradas en este estudio fueron tipo I, IIA y IIB, no observándose fibras tipo IIC, situación semejante a la observada por Essén-Gustavsson y col. (1980), Ronéus y Lindholm (1991) e Islas y col. (1996).
Los equinos mestizos de tiro presentan un mayor porcentaje de fibras tipo I y menor porcentaje de fibras tipo II comparados con los de otras razas, según los trabajos realizados por Snow y Guy (1980) y López-Rivero y col. (1990). Estos resultados son compatibles con el trabajo que efectúa el caballo de tiro, cuyo ejercicio es sostenido y a baja velocidad (Bou, 1994), para lo cual necesita un porcentaje alto de fibras tipo I en las cuales predomina el metabolismo aeróbico. Diversos estudios han demostrado que la composición de la fibra del músculo Gluteus medius en el equino (Wool y col., 1988; Essén-Gustavsson y col., 1989) presenta diferencias según la profundidad de la biopsia (Raub y col., 1985; Islas y col., 1996), situación también observada en esta investigación.
En las biopsias se observó que el porcentaje de fibras tipo I aumentó significativamente con la profundidad. Algunos investigadores sostienen que a mayor profundidad el músculo Gluteus medius realiza un metabolismo predominantemente oxidativo, permitiendo una mejor adaptación de él para funciones posturales y actividades dinámicas de bajo nivel y larga duración (Kline y Bechtel, 1988; López-Rivero y col., 1993; Islas y col., 1996).
El porcentaje de fibras tipo IIA se mantiene sin variación en las distintas profundidades estudiadas, siendo similar al caballo de tiro chileno (Islas y col., 1996) y menor al observado en el equino fina sangre (Ronéus y Lindholm, 1991) y en el caballo Andaluz, Anglo-Arabe y Arabe (López-Rivero y col., 1989). El bajo porcentaje de fibras tipo IIA encontrado en esta investigación se relacionaría con el tipo de trabajo que los equinos realizan.
Los porcentajes de las fibras tipo IIB, que se caracterizan por tener generalmente una baja capacidad oxidativa, en este estudio son menores a los encontrados en otras razas (Essén-Gustavsson y col., 1980; López-Rivero y col., 1990; Ronéus y Lindholm, 1991). Los porcentajes de fibra tipo IIB oxidativas y no oxidativas disminuyen con la profundidad de la biopsia en forma significativa. Estas fibras participan en la generación de fuerza propulsiva rápida e intensa, pero de corta duración, características fundamentales para los caballos FS de carrera (Ronéus y Lindholm, 1991), a diferencia del caballo de tiro que realiza un trabajo lento y sostenido a baja velocidad y con mayor fuerza (Bou, 1994).
El área de las fibras del equino de tiro mestizo es mayor al obtenido en otras razas en condiciones de reposo (Essén-Gustavsson y Lindholm 1985; López-Rivero y col., 1993), lo que se relacionaría con el volumen del músculo Gluteus medius que tienen estos equinos.
El diámetro mínimo obtenido en las fibras tipo I, IIA y IIB es similar al de otras razas (López-Rivero y col., 1990), no observándose diferencias con la profundidad de la biopsia. El diámetro mínimo es importante debido a su relación con la fuerza isométrica generada en la contracción y a su abastecimiento capilar. Las fibras tipo I y IIA son de menor diámetro, tienen proporcionalmente un mayor abastecimiento capilar, lo que favorece la difusión de oxígeno y sustratos al interior de las células, al mismo tiempo que facilitan la eliminación más rápida de los productos catabólicos (Essén-Gustavsson y col., 1989). Las fibras tipo IIB, de mayor diámetro, están mejor adaptadas para realizar una contracción de alta intensidad y breve duración, ideal para caballos de carrera (López-Rivero y col., 1989; López-Rivero y col., 1991).
El análisis de la actividad enzimática específica obtenida a distintas profundidades del músculo Gluteus medius muestra cambios que lo relacionan con el tipo de fibra presente en las diferentes profundidades. Así, en las biopsias más superficiales (3 cm) se observa una mayor actividad específica de la enzima LDH respecto a la actividad de las enzimas CK y CS. El músculo del equino mestizo de tiro a esa profundidad está constituido principalmente por fibras tipo IIA y IIB no oxidativas, que se caracterizan por tener un alto metabolismo glucolítico netamente anaeróbico (Kline y col., 1987; López-Rivero y col., 1992), ya que están involucradas en la generación de fuerza propulsiva de alta intensidad y de corta duración (Valberg y col., 1993).
En las biopsias de 6 cm de profundidad se observan valores de CK y LDH similares a los de la profundidad de 3 cm, lo que está relacionado con un aumento del porcentaje de las fibras tipo I y un descenso de las fibras tipo IIB. El aumento de las fibras tipo I se refleja en un incremento de la actividad de la enzima CS, lo que indica utilización del metabolismo oxidativo, propio de las fibras tipo I y IIA. Estos resultados son similares a los obtenidos por nuestro grupo en equinos de tiro criollo (Islas y col., 1996) y a los observados por López-Rivero y col. (1995) en caballos sometidos a eventos de resistencia.
En las regiones profundas del músculo (9 cm) la fibra predominante en los equinos mestizos de tiro mejorado es la tipo I, capacitada para realizar metabolismo aeróbico, utilizando sustratos extracelulares para la obtención de energía. La fibra tipo I está mejor adaptada para mantener la postura y actividades de larga duración, pero de baja intensidad (Snow y col., 1983; Bechtel y Kline, 1987), característica de los equinos de tiro analizados en este estudio, los cuales cumplen jornadas de trabajo continuo y a baja velocidad, por lo que la mayor actividad de CS a esta profundidad permitiría una mejor regulación de flujo de sustratos hacia las fibras durante ejercicios submáximos de larga duración y poca intensidad. Este mecanismo podría mejorar la eficiencia en la utilización de la energía dentro de las fibras musculares, disminuyendo el riesgo de fatiga muscular durante ejercicios de resistencia (Snow y Guy, 1981; Essén-Gustavsson y Lindholm, 1985).
Se ha demostrado que existe una correlación significativa entre el tipo de fibra y la actividad enzimática (Lindholm y Piehl, 1974; Snow, 1981; Essén-Gustavsson y col., 1984; López-Rivero y col., 1992), como se observó en este trabajo entre la fibra tipo I y la actividad de la enzima CS, lo que estaría dado por las condiciones de reposo en que se encontraban los equinos, ya que el metabolismo muscular realizado por ellos sería esencialmente aeróbico, observándose aumentos significativos de la actividad CS en relación con la profundidad de la biopsia, la cual se caracteriza por tener un elevado porcentaje de fibras tipo I, resultados similares a los obtenidos por Essén-Gustavsson (1984) y Hodgson y Rose (1987).
De los resultados obtenidos en esta investigación se puede concluir que el músculo Gluteus medius del equino mestizo de tiro descendiente del Plan de Fomento Equino presenta un metabolismo preferentemente oxidativo, expresado por una mayor proporción de fibras de contracción lenta, tipo I y una elevada actividad de la enzima CS. Estas características del músculo son consistentes con el tipo de trabajo realizado por estos animales: labores agrícolas de baja intensidad y larga duración.
RESUMEN
Se tomaron biopsias a 3, 6 y 9 cm de profundidad del músculo Gluteus medius a 16 equinos de tiro descendientes del Plan de Fomento Equino, machos y hembras entre 5 y 15 años de edad en condiciones de reposo. Las muestras fueron analizadas por las técnicas histoquímicas de miosín ATPasa para determinar la composición fibrilar, área y diámetro mínimo y la técnica NADH-TR para determinar la capacidad oxidativa de las fibras; también se determinó la actividad de las enzimas CK, LDH y CS. Los valores promedios correspondientes a los porcentajes de cada tipo de fibra fueron 33, 28, 17 y 22% para las fibras tipo I, IIA, IIB oxidativas y IIB no oxidativas, respectivamente, el área promedio fue 3.959, 4.823 y 7.087 µm2 y el diámetro mínimo fue 47, 52 y 59 µm para las fibras tipo I, IIA y IIB, respectivamente. La actividad promedio de las enzimas CK, LDH y CS fue 0.484, 0.337 y 671 µmol/min/mg de proteína, respectivamente.Los resultados obtenidos demuestran que los equinos mestizos de tiro se caracterizan porque las fibras del músculo Gluteus medius tienen una alta capacidad oxidativa. Esta característica está relacionada con el tipo de trabajo que estos animales realizan: labores agrícolas de larga duración y baja intensidad.
AGRADECIMIENTOS
Los autores desean agradecer al Fundo Pupunahue las facilidades brindadas para realizar esta investigación.
* Proyecto FONDECYT 1941005
BIBLIOGRAFIA
ALP, P., E. NEWSHOLME , V. ZAMMIT. 1976. Activities of citrate synthase and NAD-linked and NADP-linked isocitrate dehydrogenase in muscle from vertebrates and invertebrates, Biochem. J. 154 : 689-700.
BARLOW, D., J. LLOYD, P. HELLHAKE, J. SEDER. 1984. Equine muscle fiber types: A histological and histochemical analysis of selected thoroughbred yearlings, J. Equine Vet. Sci. 4: 60-66.
BECHTEL, P.J., K.H. KLINE. 1987. Muscle fiber type changes in the middle gluteal of quarter and standardbred horses from birth through one year of age. En : J.R. GILLESPIE y N.E. ROBINSON (eds.). Equine Exercise Physiology 2. Iceep Publications, Davis, California , pp. 265-270.
BOU, R. 1994. Rendimiento de trabajo y respuesta bioquímica y fisiológica del caballo de tiro sometido al ejercicio de tracción en suelo tipo trumao del área regada de Ñuble, Chile. Tesis de Grado. Universidad de Concepción, Fac. Med. Vet., Chillán, Chile.
BROOKE M.H., K.K. KAISER . 1970 . Three 'myosin ATPase' systems : The nature of their pH lability and sulphydryl dependence, J. Histochem. Cytochem. 18 : 670-672.
ESSEN, B., A. LINDHOLM, J.R. THORNTON. 1980. Histochemical properties of muscle fiber types and enzyme activities on skeletal muscles of Standardbred trotters of different ages, Equine Vet. J. 12 : 175-180.
ESSEN-GUSTAVSSON, B., K. KARLSTRÖM, A. LINDHOLM. 1984. Fibre types, enzyme activities and substrate utilisation in skeletal muscles of horses competing in endurance rides, Equine Vet. J. 16 : 197-202.
ESSEN-GUSTAVSSON, B., A. LINDHOLM. 1985. Muscle fiber characteristics of active and inactive Standardbred horses, Equine Vet. J. 17 : 434-438 .
ESSEN-GUSTAVSSON, B., D. McMIKEN, K. KARLSTRÖM, A. LINDHOLM, S. PERSSON , J. THORNTON. 1989. Muscular adaptation of horses during intensive training and detraining , Equine Vet. J. 21 : 27-33.
GALITZER, S., F. OEHME. 1985. Creatine kinase isoenzymes in bovine tissue, Am. J. Vet. Res. 46 : 1427-1429.
GUY, P.S., D.H. SNOW. 1977. The effect of training and detraining on muscle composition in the horse J. Physiol. 269 : 33-51.
HODGSON, D.R. , R.J. ROSE, J. DIMAURO, J.R. ALLEN. 1986. Effect of training on muscle composition in horses, Am. J. Vet. Res. 47 (1 ): 12-15.
HODGSON, D.R. , R.J. ROSE. 1987. Effects of a nine-month endurance training programme on muscle composition in the horse, Vet. Rec. 121 : 271-274.
ISLAS, A., J. LOPEZ-RIVERO, M. QUEZADA, G. MORA, V. AEDO, M. BRIONES, L. MARIN. 1996. Características histoquímicas de las fibras del músculo Gluteus medius en equinos de tiro, Arch. Med. Vet. 28 (1): 83-91.
KLINE, K.H., L.M. LAWRENCE, J. NOVAKOFSKY, P.J. BECHTEL. 1987. Changes in muscle fiber variation within the middle gluteal of young and mature horses as a function of sampling depth. En: J.R. GILLESPIE y N.E. ROBINSON (eds.). Equine Exercise Physiology 2. Iceep Publications, Davis, California, pp. 271-277.
KLINE, K.H., P.J. BECHTEL, 1988. Changes in the metabolic profile of the equine Gluteus medius as a function of sampling depth, Comp. Biochem. Physiol. (A) 91 : 815-819.
LEXELL, J., K. HENRIKSSON, M. SJOSTROM. 1983. Distribution of different fiber types in human skeletal muscles. A study of cross-sections of whole m. Vastus lateralis Acta Physiol. Scand. 117 : 115-122.
LINDHOLM , A. , K. PIEHL. 1974. Fibre composition, enzyme activity and concentration of metabolites and electrolytes in muscles of Standardbred horses, Acta Vet. Scand. 15 : 287-309.
LOPEZ-RIVERO, J.L., E. AGÜERA , J. VIVO , J.L. MORALES-LOPEZ, A. MARTINEZ-GALISTEO. 1989. Población y tamaño fibrilar del músculo glúteo medio de caballos pura sangre inglés, An. Anat. 35 : 19-28.
LOPEZ-RIVERO, J.L. , E. AGÜERA, J. VIVO, A. DIZ, F. MIRO. 1990. Histochemical and morphometrical study of the middle gluteal muscle in Arabian horses, J. Equine Vet. Sci. 10 : 144-148.
LOPEZ-RIVERO, J.L. , J.L. MORALES-LOPEZ , A.M. GALISTEO, E. AGÜERA . 1991. Muscle fibre type composition in untrained and endurance-trained Andalusian and Arab horses , Equine Vet. J. 23 91-93.
LOPEZ-RIVERO, J.L., E. AGÜERA , J.G. MONTERDE, J. VIVO, M.V. RODRIGUEZ-BARBUDO. 1992. Skeletal muscle fibre size in untrained and endurance-trained horses, Am. J. Vet. Res. 53 : 847-850.
LOPEZ-RIVERO, J.L., M.C. RUZ , A.L. SERRANO, A.M. GALISTEO . 1993. Efectos del entrenamiento y desentrenamiento sobre el tamaño y los tipos de fibras musculares de diferentes razas de caballos, Arch. Med. Vet. 25 : 127-136.
LOPEZ-RIVERO, J.L. 1995. Efecto del entrenamiento sobre el músculo esquelético del equino, Agro-Ciencia 11 : 71-85.
LOWRY, O., N. ROSEBROUGH , A. FARR , R. RANDALL . 1951. Protein measurements with the folling phenol reagent, J. Biol. Chem. 198 : 265-275.
PEREZ , R. , I. CABEZAS, R. GUZMAN, M. GARCIA , C. CHAVARRIA, R. SOTO . 1993. Comparación de características hipométricas de caballos mestizos descendientes de potros fina sangre y criollo de tiro , Avances Cs. Vet. 8 : 119-128.
RAUB, R.H ., P.J. BECHTEL, L.M. LAWRENCE . 1985. Variation in the distribution of the muscle fiber types in equine skeletal muscles, J. Equine Vet. Sci. 5 : 34-37.
RONEUS, M., A. LINDHOLM, A. ASHEIM . 1991. Muscle characteristics in Thoroughbreds of different ages and sexes, Equine Vet. J. 23 : 207-210.
RONEUS, M., B. ESSEN-GUSTAVSSON, A. LINDHOLM, S.G.B. PERSSON. 1992. Skeletal muscle characteristics in young trained and untrained Standardbred trotters, Equine Vet. J. 24 : 292-294.
RUDOLPH, W.G. , I.V. FONCK, P.M. SEGOVIA . 1993. Deshidrogenasa láctica sérica en el equino FSC inglés : termoinactivación de sus isoenzimas y relación con la alfa hidroxibutirato deshidrogenasa, Arch. Med. Vet 14 : 79-84.
SNOW, D.H. , P.S. GUY. 1976. Percutaneous needle muscle biopsy in the horse, J. Equine Vet. 8 : 150-155.
SNOW, D.H. , P.S. GUY . 1980 . Muscle fibre type composition of a number of limb muscles in different types of horses, Res. Vet. Sci. 28 : 137-144.
SNOW, D.H. ,
P.S. GUY. 1981. Fiber types and
enzyme activities of Gluteus medius in different breeds of horse.
En: J.B. POORTAMN
y B. NISET (eds.).
Biochemistry of Exercise IV.
Univ. Park Press, Baltimore.
SNOW, D.H. , H. RICKETTS,
D. MASON. 1983. Haematological response to racing and
training exercise in thoroughbred horses, with particular references to the
leucocyte response, Equine Vet. J. 15 : 149-154.
VALBERG, S. JÖNSSON, A. LINDHOLM, N.
HOLMGREN. 1993. Muscle histopathology
and plasma aspartate aminotransferase, creatine kinase and myoglobin changes
with exercise in horses with recurrent exertional rhabdomyolysis, Equine
Vet. J. 25 : 11-16.
WOOD , C.H. , T.T. ROSS, J.B. ARMSTRONG, D.C. HALL. 1988. Homogenety of muscle fibre composition in the M. Gluteus medius of the horse , J. Equine Vet. Sci. 8 : 294-296.
Aceptado: 09.09.96