COMUNICACIONES
Polyclonal antibodies anti-salmon immunoglobulins; potential tools for diagnosis
J.C. AGUILLON1, B.Q., PhD; P. SMITH2, M.V.,
M.Sc.; A.COLOMBO1, T.M.; M.C. MOLINA1, Q.F.;
A. FERREIRA,1 M.V., PhD.
SUMMARY
Immunoglobulins (Igs) from sera of Atlantic salmon (Salmo salar) were purified with 50% saturated ammonium sulphate followed by molecular sieving chromatography using Sephacryl S-300. The fraction of purified Igs was used as immunogen to generate a polyclonal antibody anti-salmon Igs. This antibody was affinity purified in a Sepharose-protein G column. By immunoradiometric assay against the immunogen, the rabbit IgG anti-salmon Igs showed a high specific titer (at least 1/100.000) for the antigen. This reagent offers the possibility of detection of salmon or trout (crossreactivity) humoral immune responses, both for diagnostic and research purposes.
Palabras claves: anticuerpos policlonales, salmón, diagnóstico.
Key words: polyclonal antibodies, salmon, diagnosis.
* Proyectos FONDECYT 1933-1134 y FONDEF 2-23
INTRODUCCION
En Chile la década de los 80 representó el despegue de la producción de salmón de cultivo, situándose en el segundo lugar a nivel mundial. Las especies que representan los mayores volúmenes de producción las constituyen el salmón del Atlántico (Salmo salar), salmón coho (Oncorhynchus kisutch) y trucha arco iris (Oncorhynchus mykiss). Dentro de los principales factores que ayudaron al desarrollo de esta industria en nuestro país destacó, en sus inicios, una ventajosa situación ictiosanitaria (Méndez y Munita, 1994).
Sin embargo, el éxito productivo, expresado en un aumento del retorno de divisas, se vio parcialmente obstaculizado con la aparición de bruscas mortalidades de salmón coho en el año 1989, derivadas de la infección con Piscirickettsia salmonis (Fryer y col., 1992). Esta última, actualmente, afecta al salmón del Atlántico, al salmón chinoock (Oncorhynchus tshawytscha) y trucha arco iris (Garcés y col., 1991).
Para disminuir el daño provocado por ésta y otras patologías se han desarrollado metodologías diagnósticas que apuntan a la detección del agente causal o de anticuerpos dirigidos contra éste. Dentro del primer enfoque destacan: el cultivo de microorganismos en líneas celulares (Wolf y Quimby, 1962; Fijan y col., 1983; Lannan y col., 1984), inmunofluorescencia directa o indirecta (Lewis y Allison, 1971; Lannan y col., 1991) y ensayo inmunoenzimático (ELISA) de captura del patógeno (Adams, 1992). Empleando el segundo enfoque: reacciones de aglutinación (Clark y col., 1987), inmunoprecipitación en geles (Suran y col., 1967), fijación del complemento (Dorson y Torchy, 1979) y ELISA (Kaattari y Yui, 1987).
No obstante la ventajosa sensibilidad ofrecida por los ensayos inmunométricos, su utilidad se ha visto restringida por serias dificultades, a nivel mundial, para la adquisición de anticuerpos contra inmunoglobulinas (Igs) de salmónidos. No parecería que estas dificultades se derivaran de problemas en la purificación del antígeno como se discute más adelante. Para contribuir al diagnóstico de diversas patologías, en este trabajo se describe una combinación de procedimientos experimentales simples y estándares para la producción de anticuerpos policlonales anti-Igs de salmón.
MATERIAL Y METODOS
OBTENCION DE IG DE SALMON
Obtención de suero de salmón. Un grupo de 10 salmones del Atlántico (Salmo salar) de mar, adultos, sanos, de aproximadamente 2 kg, fueron anestesiados por inmersión en una solución acuosa de benzocaína al 0.001% v/v y sangrados mediante corte transversal profundo a nivel de las branquias (arterias aorta ventral y coronaria). Los sueros fueron obtenidos por procedimientos estándares y congelados a -20°C.
Purificación de Igs de salmón. De acuerdo a lo descrito por Havarstein y col. (1988) de los sueros de salmón se aislaron sus Igs, inicialmente por medio de precipitación con sulfato de amonio al 50% de saturación. Las Igs precipitadas fueron primero lavadas con sulfato de amonio al 50% de saturación y luego redisueltas en tampón fosfato salino (PBS) para su posterior diálisis contra PBS, durante toda la noche a 4°C. Posteriormente, este material fue sometido a cromatografía de exclusión por tamaño molecular en una columna (120 x 2.5 cm) de Sephacryl S-300, eluyendo con PBS. La separación cromatográfica fue monitorizada mediante geles de poliacrilamida en presencia de dodecilsulfato de sodio (SDS-PAGE), seguida de tinción con azul de Coomassie. Aquellas fracciones que mostraron cadenas pesadas (H) y livianas (L) fueron combinadas.
GENERACION DE IGG POLICLONAL ANTI-IG DE SALMON
Inmunización de conejos. Dos conejos hembras neozelandesas, de 2 meses de edad fueron inmunizadas con 500 µg de proteína total, por inoculación, de Ig de salmón purificada (mezcla de fracciones cromatográficas 29-31). El protocolo consistió en una primera inoculación subcutánea de 5 ml (2.5 ml de coadyuvante de Freund completo y 2.5 ml de solución antigénica suspendida en suero fisiológico), una segunda subcutánea de igual volumen (2.5 ml de coadyuvante de Freund incompleto y 2.5 ml de solución antigénica) y una última intraperitoneal de 1 ml (0.5 ml de coadyuvante Freund incompleto y 0.5 ml de solución antigénica), todas ellas separadas por una semana. Las sangrías fueron semanalmente, empezando con una preinmune. Los sueros se obtuvieron mediante procedimientos estándares.
Purificación por afinidad de IgG de conejo anti-Ig de salmón. Alícuotas de 5 ml de suero de conejo (tercera sangría) fueron pasadas a través de una columna de afinidad de Sefarosa-proteína G. La columna, con la mayor parte de las IgG retenidas, fue lavada con 100 veces su volumen de PBS. Luego ésta fue eluida con buffer glicina (glicina 0.1M / NaCl 0.15M, pH 2.6), colectando el material eluido sobre buffer Tris 2M pH 8.0. La pureza de la IgG de conejo fue analizada mediante SDS-PAGE, seguida de tinción con azul de Coomassie.
Titulación de la IgG anti-Ig de salmón. Pocillos de placas de cloruro de polivinilo (PVC) fueron sensibilizados con 50 µl de una solución 20 µg/ml de Ig de salmón purificada. Los sitios reactivos remanentes se bloquearon con albúmina sérica bovina al 1% p/v en PBS. Diluciones seriadas de la IgG anti-Ig de salmón se hicieron reaccionar con la Ig adsorbida a la fase sólida. El revelado se efectuó con IgG de cabra anti-IgG de conejo, purificada por afinidad y radiomarcada con 125I. La radiactividad asociada específicamente a los pocillos fue medida en un contador gamma.
RESULTADOS Y DISCUSION
Aunque en los mamíferos hay cinco clases o isotipos de Igs, en los peces teleósteos sólo se ha demostrado la existencia de una. Se trata de una macroglobulina similar a la IgM, tetramérica, compuesta de una estructura básica de dos cadenas H y dos L. Dependiendo de las especies de peces, su masa molecular varía entre 600 y 800 kDa (Clem, 1971).
Para la purificación de las Igs de peces se ha descrito una variedad de técnicas, dentro de las que destacan la precipitación con sulfato de amonio saturado, separación por cromatografía de exclusión por tamaño molecular y por intercambio iónico (Acton y col., 1971; Havarstein y col., 1988). En este trabajo, para la purificación de las Igs de salmón, se usó una combinación de las dos primeras.
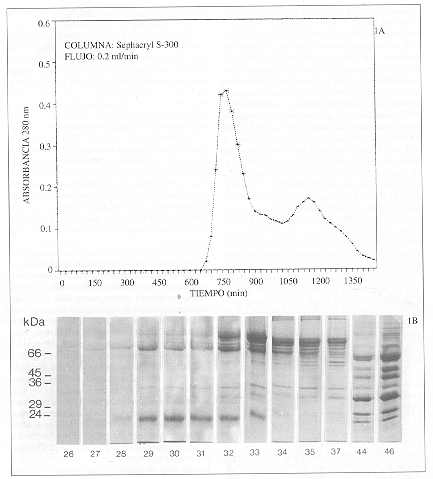
En la figura 1A se muestra el perfil de separación, para las Igs de salmón, obtenido al pasar el producto fraccionado con sulfato de amonio al 50% de saturación a través de una columna de exclusión por tamaño molecular de Sephacryl S-300. De los dos picos resueltos, el primero contiene a las Igs y, el segundo, a otros componentes proteicos y remanentes de albúmina sérica. Esto se deduce al seguir la purificación mediante SDS-PAGE seguida de tinción con azul de Coomassie, para las distintas fracciones colectadas (fig. 1B) y al comparar sus masas moleculares relativas con la de los estándares. Las Igs fueron colectadas, principalmente, entre las fracciones 29 y 31 (fig. 1B, carriles 29-31), observándose también su presencia en las fracciones 27, 28, 32 y 33. Las cadenas H y L (fig. 1B, carriles 20-32) alcanzaron una migración electroforética aproximada, correspondiente a los 70 y 25 kDa, respectivamente, valores descritos para esta Ig (Havarstein y col., 1988). En las fracciones más enriquecidas de Igs de salmón (29-31), éstas representan más del 80% de las proteínas totales.
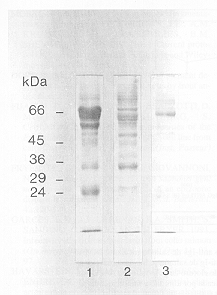
En la figura 2 se muestra el grado de purificación alcanzado con la precipitación con sulfato de amonio (carril 2) y luego al usar cromatografía de exclusión por tamaño molecular (carril 3). El sulfato de amonio eliminó la mayor parte de la albúmina sérica, y la separación cromatográfica generó un producto adecuado para obtención de un anticuerpo policlonal.
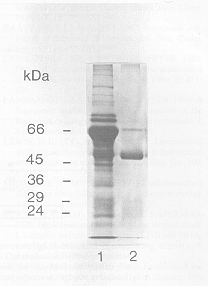
Los anticuerpos anti-Igs de salmón fueron aislados desde el suero completo del conejo inmune mediante cromatografía de afinidad, utilizando proteína G, la cual tiene afinidad por el dominio Cg2 de la IgG (Coligan y col., 1995). En la figura 3 se puede apreciar la pureza de estos anticuerpos, antes (carril 1) y después (carril 2) de la separación cromatográfica. Se distingue claramente la cadena H, alrededor de los 50 kDa, mientras la cadena L se visualiza débilmente, debido a una baja captación del colorante, derivado del alto grado de glicosilación de esta cadena. La presencia de una ligera banda alrededor de los 66 kDa (carril 2) representa un pequeño porcentaje (no más de 10%) de albúmina sérica de conejo que permaneció adsorbida a la proteína G. En un solo paso se logró obtener más de un 90% de pureza.
El anticuerpo purificado mantuvo su capacidad de combinarse con el antígeno (Igs de salmón), verificado mediante ensayo inmunorradiométrico (IRMA) en diluciones de por lo menos 1/100.000 (fig. 4).
Mediante inmunoelectrotransferencia, tanto contra suero completo como contra Igs purificadas, provenientes de salmón del Atlántico, el anticuerpo reaccionó predominantemente con las cadenas pesadas y livianas de las Igs. En el primer caso, la intensidad del reconocimiento fue más débil, lo cual se explica por la menor concentración relativa de las Igs en el suero completo de salmón (resultados no mostrados).
Adicionalmente, mediante IRMA, este anticuerpo demostró buena reactividad cruzada con las Igs de salmón coho y trucha arco iris. Desde el punto de vista diagnóstico esto es ventajoso, dado que la utilización de un solo reactivo permitiría detectar infecciones en diversas especies de salmónidos. La técnica de IRMA es equivalente a la de ELISA ya que sólo cambia la naturaleza de la inmunosonda (radiactiva versus enzimática).
La generación de anticuerpos de esta naturaleza no parece trivial. La etapa de purificación de las Igs de salmónidos es un paso previo ineludible. El reactivo generado puede ser útil en el análisis diagnóstico e investigación. Así, podría facilitar la detección de anticuerpos dirigidos contra un patógeno, ya sea viral, bacteriano, e incluso parasitario, mediante técnicas de inmunofluorescencia, ensayos inmunométricos, inmunodot e inmunoelectrotransferencia. También podría permitir adquirir información relacionada con la inmunidad humoral en peces salmónidos, frente al desafío con patógenos definidos.
El enfoque experimental sería usar el anticuerpo de conejo anti-Igs de salmón como inmunosonda de detección de estas últimas, revelando con otro anticuerpo dirigido contra el isotipo de la especie (conejo) en que fue generado. Otro enfoque sería usar directamente el anticuerpo, purificado por afinidad por el antígeno, como inmunosonda de detección de las Igs de salmón, unidas al antígeno del patógeno adsorbido a una fase sólida como PVC.
Alternativamente, la unión de la IgG de conejo a las Igs de salmón podría detectarse con proteína A o IgG de cabra anti-IgG de conejo, ambas transformadas en inmunosondas enzimáticas o radiactivas.
Figura 1-A. Purificación de inmunoglobulinas
(Igs) de salmón mediante cromatografía por tamaño molecular,
utilizando Sephacryl S-300. Las Igs, previamente precipitadas con sulfato
de amonio al 50% de saturación, eluyeron en el primer pico (fig. 1B,
fracciones 29-31). 1B. Análisis electroforético de las fracciones
de Igs de salmón purificadas por tamaño molecular. Las fracciones
cromatográficas (26-35, 37, 44 y 46) fueron separadas mediante SDS-PAGE,
seguida de tinción con azul de Coomassie. Las Igs eluyeron principalmente
entre las fracciones 29 y 31. En el gel se cargaron 30 µl de cada una
de las fracciones en estudio. Los números de la izquierda representan
estándares de masa molecular.
1A. Purification of salmon immunoglobulins (Igs) by molecular sieving chromatography
using Sephacryl S-300. Igs, previously precipitated with 50% saturated ammonium
sulphate, eluted in the first peak (Figure 1B, fractions 29-31). 1B. Electrophoretic
analysis of the purified salmon Ig fractions by molecular sieving. Chromatographic
fractions (26-35, 37, 44 and 46) were separated by SDS-PAGE and stained with
Coomassie blue. Igs eluted mainly in fractions 29 and 31. 30 µl of each
fraction were run in the gel. Numbers to the left represent molecular mass
standards.
 |
 |
|
Figura 2. Análisis
electroforético de la purificación de inmunoglobulinas (Igs)
de salmón. Las muestras se separaron mediante SDS-PAGE seguida
de tinción con azul de Coomassie. Carril 1: suero completo de salmón
del Atlántico (3 µl). Carril 2: Igs de salmón precipitadas
con sulfato de amonio al 50% de saturación (1 µl). Carril
3: Igs de salmón precipitadas con sulfato de amonio y purificadas
por cromatografía de tamaño molecular con Sephacryl S-300
(20 µl). Los números de la izquierda representan estándares
de masa molecular.
Electrophoretic analysis of salmon immunoglobulin (Ig) purification. Samples were separated by SDS-PAGE and stained with Coomassie blue. Track 1: complete salmon serum (3 µl). Track 2: salmon Igs, precipitated with 50% saturated ammonium sulphate (1µl). Track 3: salmon Igs, precipitated with ammonium sulphate and purified by molecular sieving chromatography using Sephacryl S-300 (20 µl). Numbers to the left represent molecular mass standards. |
Figura 3. Análisis
electroforético de IgG de conejo anti-Igs de salmón, purificada
por afinidad, seguido de tinción con azul de Coomassie. El suero
completo de conejo fue purificado a través de una columna de Sefarosa-proteína
G. Carril l: 1 µl de suero completo de conejo anti-Igs de salmón.
Carril 2: IgG de conejo anti-Igs de salmón purificada por afinidad.
Los números de la izquierda representan estándares de masa
molecular.
Electrophoretic analysis of rabbit anti-salmon Igs IgG, affinity purified and stained with Coomassie blue. The whole rabbit serum was purified by a Sepharose-protein G column. Track 1: whole rabbit serum anti-salmon Igs (1 µl). Track 2: rabbit anti-salmon Igs IgG, affinity purified. Numbers to the left represent molecular mass standards. |
Finalmente, debe destacarse que comercialmente sólo se dispone de un anticuerpo policlonal contra Igs de trucha (generado en cabra por Kirkegaard & Perry Laboratories, Inc., U.S.A.). Consultada la literatura no hemos detectado publicaciones que describan el procedimiento empleado para generar este reactivo. Además, su accesibilidad a los productores nacionales es limitada al no existir representante regional para adquirir el producto. Por otra parte, la compañía BIOS Chile. I.G.S.A. generó en ratones un anticuerpo policlonal contra Igs de salmón, sin embargo, actualmente no se encuentra disponible. Por último, el uso de anticuerpos monoclonales murinos está restringido al anticuerpo 4C10, contra salmón del Atlántico. Este no se produce comercialmente, sólo se puede obtener por donación del Instituto de Aquacultura de la Universidad de Stirling, Escocia, por lo cual su uso se restringe a algunos laboratorios de investigación.
Los hechos anteriores señalan, entonces, la conveniencia de contar con un procedimiento simple para generar el reactivo en un país cuya producción de salmónidos es de las mayores a nivel mundial.
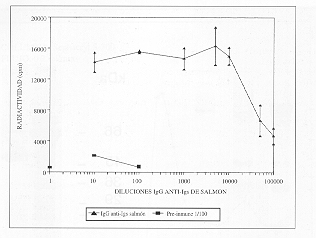
Figura 4. Titulación de
IgG de conejo anti-Igs de salmón mediante IRMA.
Diluciones, por triplicado, de la IgG anti-Ig de salmón se hicieron
reaccionar
con Ig de salmón adsorbida a pocillos de cloruro de polivinilo. Se
reveló con
IgG de cabra anti-IgG de conejo, purificada por afinidad y radiomarcada.
Rabbit anti-salmon Igs IgG titration
by IRMA. Dilutions, in triplicates, of
nti-salmon Igs IgG were tested with salmon Igs adsorbed to wells of polyvinyl
chloride microtitration plates. Detection was with goat IgG anti-rabbit IgG,
affinity purified and radiolabelled.
RESUMEN
A partir de sueros de salmón del Atlántico (Salmo salar) se han purificado sus inmunoglobulinas (Igs) mediante precipitación con sulfato de amonio al 50% de saturación, seguido de cromatografía de exclusión por tamaño molecular en Sephacryl S-300. La fracción de Igs purificadas fue utilizada como inmunógeno para generar un anticuerpo policlonal anti-Igs de salmón, el cual fue purificado por afinidad usando Sefarosa-proteína G. Mediante ensayo inmunorradiométrico contra el inmunógeno, la IgG anti-Igs de salmón demostró un elevado título (a lo menos 1/100.000) específico para el antígeno. El anticuerpo generado ofrece la posibilidad de detectar respuestas humorales en salmón y trucha (reactividad cruzada), con fines diagnósticos o de investigación.
BIBLIOGRAFIA
ACTON, R.T., P.F. WEINHEIMER, S.J. HALL, W. NIEDERMEIER, E.SHELTON, J.C. BENNETT. 1971. Tetrameric immune macroglobulins in three orders of bony fishes, Proc. Natl. Acad. Sci. USA 68: 107-111.
ADAMS, A. 1992. Sandwich enzyme linked immunosorbent assay (ELISA) to detect and quantify bacterial pathogens in fish tissue. En: J.S. STOLEN, T.C. FLETCHER, D.P. ANDERSON, S.L. KAATTARI, A.F. ROWLEY(eds.) Techniques in fish immunology. SOS Publications, Fair Haven, New Jersey.
CLARK, T.G., H.W. DICKERSON, J.B. GRATZEK, R.C. FINDLY. 1987. In vitro response of Ichthyophthirius multifiliis to sera from immune channel catfish, J. Fish Biol. 31 (supplement A): 203-208.
CLEM, L.W. 1971. Phylogeny of immunoglobulin structure and function. IV. Immunoglobulins of the giant grouper, Epinephelusitaira, J. Biol. Chem. 246: 9-15.
MORRISON, S.L. 1995. Purification and fragmentation of antibodies. En: COLIGAN, J.E., A.M. KRUISBEEK, D.H. MARGULIES, E.M. SHEVACH, W. STROBER (eds.). Current protocols in immunology. Greene Publishing and Wiley-Interscience, New York.
DORSON, M., C. TORCHY. 1979. Complement dependent neutralization of Egtved virus by trout antibodies, J. Fish Dis. 2: 345-347.
FIJAN, N., D. SULIMANOVIC, M. BEARZOTTI, D. MUZINIC, L.O. ZWILLENBERG, S. CHILMONCZYK. 1983. Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio, Ann. Virol. (Inst. Pasteur) 134E : 207-220.
FRYER, J.L., C.N. LANNAN, S.G. GIOVANNONI, N.D. WOOD. 1992. Piscirickettsia salmonis gen. nov., sp. nov., the causative agent of an epizootic disease in salmonid fishes, Int. J. Syst. Bacteriol. 42: 120-126.
GARCES, L.H., J.J. LARENAS, P.A. SMITH, S. SANDINO, C.N. LANNAN, J.L. FRYER. 1991. Infectivity of a rickettsia isolated from coho salmon (Oncorhynchus kisutch), Dis. Aquat. Org. 11: 93-97.
HAVARSTEIN, L.S., P.M. ASSJORD, S. NESS, C. ENDRESEN. 1988. Purification and partial characterization of an IgM-like serum immunoglobulin from Atlantic salmon (Salmo salar), Dev. Comp. Immunol.12: 773-785.
KAATTARI, S.L., M.A. YUI. 1987. Polyclonal activation of salmonid B lymphocytes, Dev. Comp. Immunol. 11: 155-165.
LANNAN, C.N., J.R. WINTON, J.L. FRYER. 1984. Fish cell lines. establishment and characterization of nine cell lines from salmonids, In Vitro 20: 671-676.
LANNAN, C.N., S.A. EWING, J.L. FRYER. 1991. A fluorescent antibody test for the detection of the Rickettsia causing disease in Chilean salmonids, J. Aquat. Anim. Health 4: 229-234.
LEWIS, D.H., T.C. ALLISON. 1971. Detection of antibodies to Aeromonas liquifaciens in fish utilized in lunar exposure studies, Trans. Am. Fish. Soc. 100: 575-578.
MENDEZ, R., C. MUNITA. 1994. La salmonicultura chilena en 1993. Balance de una creciente actividad, Aquanoticias Internacional 20: 24-39.
SURAN A.A., M.H. TARAIL, B.W. PAPERMASTER. 1967. Immunoglobulins of the leopard shark. I. Isolation and characterization of 17S and 7S immunoglobulins with precipitating activity, J. Immunol. 99: 679-686.
WOLF K., M.C. QUIMBY. 1962. Established eurythermic line of fish cells in vitro. Science 135: 1065-1066.
Aceptado: 15.07.96