R. PEREZ, M.V., M.Sc.; M. GARCIA, M.V.; I. CABEZAS, M.V.; R. GUZMAN, M.V.; V. MERINO, BQ. M.Sc.; S. VALENZUELA, BQ.; C. GONZALEZ, Lic. Cs. Vet.
Laboratorio de Fisiología del Ejercicio. Departamento de Ciencias Clínicas. Facultad de Medicina Veterinaria. Universidad de Concepción, Campus Chillán, Casilla 537, Chillán, Chile.
SUMMARY
Physical activity, cardiovascular and biochemical changes of Chilean purebred horses to rodeo competitions
A study, in order to characterize the level of physical work and the physiological and biochemical response of the Chilean purebred horse during the rodeo competition was carried out. Hippometric measurements of horses were taken and the weight of the rider, saddle and bridle were also determined. The perimeter of the rodeo track was measured in order to determine the work speed during the competition. Before and at different times after the first ride of a steer, the heart rate was recorded and blood samples were taken to measure packed cell volume (PCV), total proteins, haemoglobin, glucose, lactic acid, triglycerides, insulin, cortisol and the electrolytes: sodium, potassium, calcium and chloride. The plasma activity of the enzymes creatinekinase (CK), lactate dehydrogenase (LDH) and aspartate aminotransferase (AST) were also determined.
It is concluded that the Chilean purebred horse is characterized by a low height of withers, with a body length/heart girth ratio that agrees with a medium size horse of 392 ± 33 kg body weight. These characteristics match those described in the standard values for the Chilean criollo horse. The principal physical activity that the Chilean horse carries out during the rodeo is to ride a 318 ± 11 kg body weight steer and to stop it in the track wall, carrying on its back a weight equivalent to 24% of its body weight, at speeds ranging between 5-8 m/s over a sand and circular track; exercise that determines significant increases in HR, PCV and in the plasma concentrations of haemoglobin, glucose, lactic acid, triglycerides, insulin, cortisol and in the serum activity of enzymes CK, LDH and AST. Increases in total serum proteins and a delayed decrease in serum potassium concentration were the most significant changes observed in the hydrosaline balance. It is concluded that for Chilean horses, the rodeo represents an exercise of force, speed and resistance which induces changes in their cardiovascular, hydrosaline, metabolic and endocrine activity, responses which are similar to those observed in high intensity exercises.
Palabras claves: caballos, rodeo, ejercicio, fisiología.
Key words: horses, rodeo, exercise, physiology.
INTRODUCCION
En Chile el rodeo representa un deporte nacional de car?cter tradicional y masivo, motivo por el cual se ha constituido en el principal estímulo para la crianza del caballo de "Raza Criolla Chilena" dado el auge que ha tenido al ser considerado como actividad deportiva reconocida por la Dirección General de Deportes y Recreación (Pinochet, 1980).
Un requisito importante para que un caballo pueda participar en los rodeos oficiales es el de estar inscrito en el "Registro Genealógico de la Raza Criolla Chilena", para lo cual debe tener una conformación que le permita cumplir con el estándar de la raza. Morfológicamente debe poseer un perfil convexo o rectilíneo, lomo firme, grupa inclinada, pecho ancho y musculoso, cola gruesa y abundante, caña corta y cascos pequeños ((Pinochet, 1980); Valdivia, 1995). Su tipo es de un caballo musculoso, rápido y ágil en sus movimientos, debe tener además un sello racial de conjunto que completen el estándar zootécnico dirigido a un ejemplar de buenas aptitudes de silla y condiciones vaqueras (Pinochet, 1980).
La capacidad atlética de un individuo refleja la eficiencia para lograr la velocidad deseada y/o resistencia requerida para realizar un trabajo determinado y depende de los efectos combinados de factores genéticos y ambientales como es el entrenamiento (Cardinet, 1989). Al ser seleccionado en el transcurso de los años para desarrollar un determinado trabajo muscular, el caballo chileno se ha especializado en su habilidad y destreza para las labores del rodeo. Con el transcurso de las generaciones gran parte de su capacidad competitiva se ha ido transmitiendo a la descendencia mediante cruzamientos dirigidos. Sin embargo, a pesar de la similitud genética entre los componentes de la raza, al igual que en los caballos de carrera, persisten diferencias individuales que hacen que sólo algunos individuos dentro de la población presenten mejores aptitudes y rendimientos competitivos.
En las últimas décadas ha habido un considerable avance en el conocimiento científico acerca de la adaptación fisiológica del atleta equino al ejercicio (Martínez, 1989). Son numerosos y variados los trabajos en los cuales ha sido posible caracterizar los cambios fisiológicos y bioquímicos observados en los caballos posterior a competencias de velocidad (Evans y Rose, 1987; Evans y col., 1990), cabalgatas de resistencia (Cardinet y col., 1963; Deldar y col, 1982; Rose, 1986), competencias de polo (Craig y col., 1985), de trote (Krzywanek, 1974) y salto (Lekeux y col., 1991). Los resultados de estos estudios han demostrado que durante el ejercicio se producen diversos cambios en la composición y distribución de los constituyentes del plasma que reflejan el nivel de adaptación cardiorrespiratoria y metabólica destinados a lograr un adecuado aporte de oxígeno a los tejidos, favorecer la remoción de los productos metabólicos de desecho y facilitar la pérdida de calor generado por el aumento de la actividad muscular (Deldar y col., 1982; Snow y col., 1982; Rose, 1986). Estos cambios guardan una estrecha relación con la intensidad del trabajo realizado, la velocidad desarrollada, el tiempo de ejercicio y el nivel de entrenamiento (Krzywanek y col., 1976; Milne y col., 1976; Snow y col., 1982; Martínez y col., 1987). Es por ello que el conocimiento adecuado de los fenómenos involucrados en la adaptación fisiológica y bioquímica al ejercicio puede proveer las bases para mejorar el manejo, los programas de entrenamiento, la evaluación de la performance potencial y facilitar la comprensión de los procesos patológicos derivados del ejercicio (Lekeux y col., 1991).
A pesar del amplio interés deportivo del rodeo y el alto valor económico de la mayoría de los ejemplares que participan en las diferentes competencias organizadas a lo largo del país por la Asociación Nacional de Rodeo, existen muy pocos antecedentes bibliográficos y no hay estudios científicos que entreguen información acerca de la capacidad atlética y el grado de adaptación fisiológica y bioquímica que presentan estos caballos durante las competencias. Un adecuado conocimiento de los cambios fisiológicos y bioquímicos que ocurren como resultado del esfuerzo muscular que realizan los caballos chilenos durante el rodeo puede contribuir a establecer las bases objetivas para mejorar las normas de manejo, los programas de entrenamiento y evaluar el potencial rendimiento del caballo. Además, puede permitir a los médicos veterinarios contar con elementos objetivos para evaluar la condición clínica de los caballos de rodeo sobre todo en condiciones de sobreesfuerzo y fatiga muscular.
El presente trabajo tiene por objetivo hacer una contribución al conocimiento de la fisiología del ejercicio del caballo fina sangre chileno y estuvo dirigido a caracterizar el nivel de esfuerzo físico y los cambios fisiológicos y bioquímicos que se observan en estos caballos en respuesta a la competencia de rodeo.
MATERIAL Y METODOS
El estudio se realizó en caballos fina sangre chilenos inscritos, que concurrieron a los rodeos oficiales correspondiente a la temporada 1995-1996 de la ciudades de Chillán, Pinto, San Ignacio, San Carlos y Parral.
Para determinar el nivel de esfuerzo que realizan los caballos chilenos en el rodeo se determinaron sus medidas hipométricas, se pesaron el jinete y los aperos, se cronometró el tiempo de corrida en el apiñadero y en la cancha (pista de corrida). Ademas, se midió la frecuencia cardíaca y se extrajeron muestras de sangre para determinar algunas variables hematológicas y bioquímicas.
a) Hipometría. Se midió un total de 152 caballos, de los cuales 51 fueron yeguas, 63 caballos y 38 potros, en los que se determinaron sus medidas hipométricas de alzada (A), perímetro torácico (P.T.), longitud escapuloisquial (L.E.I.) y perímetros metacarpiano (P.M.), según método descrito anteriormente en caballos de tiro (Pérez y col., 1993). Para ello, los caballos se ubicaron en una superficie plana horizontal con la cabeza erguida en posición natural, con sus extremidades paralelas entre sí y perpendiculares respecto del plano de sustentación. El peso de los caballos se determinó mediante una balanza electrónica digital (Ruddweigh), para lo cual se habilitó una plataforma de madera que fue colocada sobre los sensores de la balanza. Los animales se colocaron sobre esta plataforma con sus 4 miembros en paralelo y se mantuvieron durante 30 segundos o hasta que la balanza diera una lectura estable.
La edad se determinó considerando la marca sobre la región escapular izquierda que indica el año de inscripción de nacimiento en el Registro Genealógico de la Raza Criolla Chilena. En caso de duda se recurrió a la determinación de edad según cronometría dentaria (Ensminger, 1973).
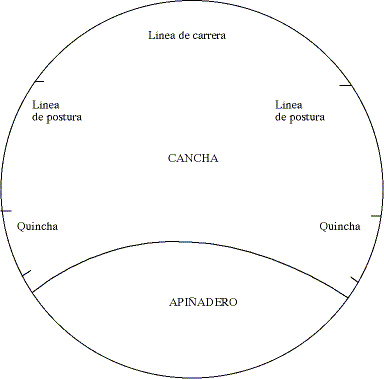
b) Caracterización del esfuerzo físico que realiza el caballo chileno durante el rodeo. Previo a cada rodeo se midió el perímetro del apiñadero y de la cancha en sus diferentes porciones (quincha, líneas de postura y línea de carrera). Un esquema de la medialuna con sus diferentes porciones se muestra en la figura 1. Cada uno de los segmentos de la cancha aparece claramente delimitado mediante líneas verticales sobre las paredes de la medialuna.
Durante la corrida en la medialuna se cronometró el tiempo de corrida en el apiñadero, el que correspondió al lapso transcurrido entre la entrada del animal al apiñadero y la salida de éste a la cancha luego de transcurridas 2,5 vueltas en su interior. Debido a que el largo de la quincha es de 12 metros y la atajada del novillo se puede producir en cualquier porción de ella, la medición del tiempo de corrida en la cancha se estandarizó a la línea de carrera, que en la mayoría de las medialunas tuvo un arco de 28 metros. Cada corrida de novillo en la cancha estuvo compuesta de 3 atajadas, por lo tanto se cronometraron 3 tiempos de carrera más un tiempo adicional para la entrega del novillo a los corrales. Luego de conocidos los valores de distancia recorrida en la línea de carrera, se dividieron por el tiempo que demora el paso del novillo por este segmento con el fin de determinar la velocidad de carrera, para cada una de las corridas consideradas en el rodeo.
Figura 1. Diagrama que muestra los diferentes segmentos de la medialuna.
Diagram with the different segments of the rodeo track.
Paralelo a la determinación de las medidas hipométricas de cada caballo se pesaron sus respectivos jinetes y monturas con el fin de determinar la carga real al dorso que lleva cada caballo durante la corrida. Además, en cada rodeo se pesaron 10 novillos elegidos al azar a los cuales también se les midió la alzada.
c) Determinación de variables fisiológicas y bioquímicas. La frecuencia cardíaca se determinó mediante la auscultación directa con fonendoscopio de la región látero-cordial izquierda.
MUESTRAS DE SANGRE. Se extrajo un total de 141 muestras, de las cuales 71 fueron obtenidas en reposo, entre las 7.15 y las 10.00 de la mañana, previo a la colocación de montura y aperos. El resto de las muestras se obtuvieron entre los 5-15 minutos posteriores a la corrida del primer animal, según las facilidades otorgadas por los corredores. Otras muestras fueron extraídas a las 8 y 24 horas de iniciada la participación de los caballos en el primer día de rodeo. Por lo tanto, la muestra de 24 h se tomó en la mañana del segundo día de rodeo, en condiciones de reposo similares a las descritas anteriormente.
De cada caballo se extrajeron 18 ml de sangre por punción directa de la vena yugular, con una jeringa estéril en cada tiempo de muestreo. La sangre extraída se repartió en tres tubos de la siguiente forma:
-Tres mililitros se colocaron en un tubo que contenía heparina y fluoruro de sodio, como anticoagulante y bloqueador de la glicólisis respectivamente, para extraer el plasma y medir las concentraciones de glucosa y ácido láctico.
-Ocho mililitros se pusieron en un tubo con heparina para medir posteriormente concentración de hemoglobina y volumen globular en sangre entera y en el plasma determinar proteínas plasmáticas totales, insulina, cortisol y la actividad de las enzimas creatinaquinasa (CK), aspartato aminotransferasa (AST) y deshidrogenasa láctica (LDH). Las muestras se mantuvieron en frío mediante un termo con hielo hasta el arribo al laboratorio.
En un tercer tubo sin anticoagulante se colocaron 7 ml de sangre, el que se mantuvo a temperatura ambiente para facilitar la coagulación con el fin de extraer el suero y determinar las concentraciones de triglicéridos y electrólitos (sodio, potasio, cloruro y calcio).
Las muestras fueron centrifugadas a 2.500 rpm por 20 minutos, para obtener el plasma o suero, que se colocó en tubos Eppendorf y se mantuvieron en refrigeración a 4º C hasta su análisis, que se realizó dentro de las 24 h posteriores a su recolección.
La concentración de hemoglobina se determinó mediante el método de la cianometahemoglobina (Boehringer, Mannheim ) y las lecturas se realizaron a una longitud de onda de 546 nm en un espectrofotómetro (Spectronic 601). El volumen globular se determinó por el método de microhematocrito leyendo directamente en la escala de la centrífuga de microhematocrito (IEC-MB Centrifuge).
Las muestras de plasma para la determinación de glucosa y ácido láctico fueron desproteinizadas con ácido perclórico 0.6M y luego centrifugadas a 10.000 rpm por 10 minutos para extraer el sobrenadante. La concentración de glucosa se determinó mediante el método enzimático de la glucosa oxidasa peroxidasa (Boehringer, Mannheim) a una longitud de onda de 546 nm. La concentración de ácido láctico se determinó a través del método enzimático de la lactato deshidrogenasa/NAD (Boehringer, Mannheim) a una longitud de onda de 365 nm.
La concentración de proteínas plasmáticas totales se determinó mediante el método colorimétrico de Biuret (Boehringer, Mannheim) y se leyó a una longitud de onda de 546 nm.
La concentración de triglicéridos se determinó a través del método enzimático colorimétrico (Boehringer, Mannheim), que consiste en la hidrólisis enzimática de los triglicéridos y determinación subsiguiente del glicerol formado; la lectura se hizo a una longitud de onda de 500 nm.
Se determinó la actividad de las enzimas creatinaquinasa (CK; E.C. 2.7.3.2), aspartato aminotransferasa (AST; E.C. 2.6.1.1) y lactato deshidrogenasa (LDH; E.C. 1.1.1.27), mediante métodos cinéticos (UV 365 nm), a una temperatura de 25º C; la lectura se realizó en un espectrofotómetro (Spectronic 601) con celdilla termorregulada, utilizando reactivos comerciales (Boehringer, Mannheim). La actividad se expresó en U/L.
CORTISOL E INSULINA. Se determinaron por radioinmunoanálisis (RIA), utilizando reactivos comerciales (Coat-A-Count, Diagnostic Products Corporation, Los Angeles. CA). La lectura de la reacción se realizó en un contador de centelleo gamma (1271 Riagamma, LKB, Wallac).
Las concentraciones de calcio, potasio y sodio plasmáticos se determinaron en un fotómetro de llama (Jenway, modelo PF7). Mientras que la concentración de cloruros se determinó en un medidor de cloruros (Jenway, modelo PCLM3). Los resultados se expresaron en mEq/L.
DISEÑO EXPERIMENTAL. Debido a la imposibilidad de hacer muestreos sucesivos de los caballos durante el rodeo y según las facilidades otorgadas por los corredores para la extracción de sangre, los resultados se distribuyeron en 4 grupos experimentales según el tiempo de muestreo, de acuerdo al siguente esquema:
Grupo I: En reposo, en la mañana, previo a la colocación de montura y aperos (n = 71).
Grupo II: 5 a 15 minutos posteriores a la corrida del primer animal (n = 40).
Grupo III: 8 horas posteriores a la corrida del primer animal (n = 12).
Grupo IV: 24 horas posteriores a la corrida del primer animal (n = 19). Los caballos considerados en los grupos II, III y IV correspondieron sólo a los que se les habían extraído muestras en reposo. En estas condiciones sólo se consideraron aquellos caballos que tuvieran sus valores de reposo dentro de rangos normales (Rose y Hodgson, 1993).
ANALISIS ESTADISTICO. Los resultados se expresaron como promedio ± desviación estándar ( ± DE) y las diferencias entre las medias se compararon mediante un análisis de varianza (ANDEVA) asociado a una prueba de comparaciones múltiples de Newman-Keuls, con un nivel de significancia de p<0.05.
RESULTADOS
HIPOMETRIA. Los caballos considerados en el estudio tenían una edad promedio de 9.8 ± 3.6 años y presentaron promedios de 139.9 ± 3.7 cm de alzada, 169.7 ± 6.4 cm de perímetro torácico, 147.4 ± 7.1 cm de longitud escapuloisquial y 19.4 ± 0.9 cm de perímetro metacarpiano. El peso corporal promedio fue de 392.1 ± 33.8 kg.
ESFUERZO FISICO. El peso promedio de los jinetes fue de 79.6 ± 10.0 kg, lo que sumado al peso de montura y riendas (13.3 ± 0.4) da un peso total de 92.9 ± 10.0 kg, valor que representa el 23.7 ± 3.7% del peso corporal de los caballos. El promedio de peso de los novillos fue de 318.4 ± 11.0 kg, los que presentaron una alzada promedio de 118 ± 2.1 cm.
La velocidad desarrollada por los caballos durante la corrida del novillo en el apiñadero fue de 6.95 ± 1.17 m/s, promedios que fluctúan entre 6.40 ± 1.31 m/s en el rodeo de Parral y los 7.53 ± 0.59 m/s en el rodeo de Chillán (cuadro 1).
La mayor velocidad desarrollada por los caballos se observó en la primera corrida del novillo, donde se lograron valores de 7.95 ± 1.13 m/s, promedio que va decreciendo a 6.27 ± 1.17 m/s en la segunda corrida y a 4.98 ± 0.98 m/s en la tercera. La vuelta después de la tercera atajada para entregar el novillo se realizó a una velocidad promedio de 3.18 ± 1.16 m/s (cuadro 1). La corrida completa del novillo en la medialuna tuvo una duración que fluctuó entre 96 a 130 segundos.
VARIABLES FISIOLOGICAS Y BIOQUIMICAS. La figura 2 muestra las variaciones de la frecuencia cardíaca observadas en los caballos previo y posterior al rodeo. Como se observa, la F.C. aumenta desde un valor de reposo de 42.0 ± 1.0 lat/min a valores de 110.0 ± 4.0 lat/min y de 62.0 ± 3.0 lat/min a los 5 y 15 min postejercicio. A las 6 y 24 h postejercicio la F.C. mantiene promedios similares al reposo.
En el cuadro 2 se muestran los valores promedios de las variables hematológicas y bioquímicas observadas en los caballos chilenos previo y posterior al rodeo. A los 15 minutos postejercicio se observaron incrementos significativos en los promedios de volumen globular y en las concentraciones de hemoglobina, proteínas totales, glucosa, ácido láctico y triglicéridos. También se observaron aumentos significativos en las concentraciones de insulina y cortisol. De igual modo se observaron incrementos significativos en los niveles de actividad plasmática de CK, LDH y AST. Sin embargo, los incrementos en la actividad de las enzimas estudiadas, presentaron patrones de variación que fueron diferentes entre ellas, así por ejemplo los niveles de CK y LDH aumentaron en forma significativa (p < 0.05) en los tiempos inmediatamente posteriores al ejercicio (15 min).
En cambio, AST presentó aumentos significativos (p < 0.05) a las 24 h postejercicio.
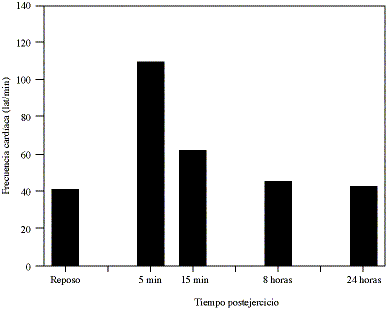
Figura 2. Promedios de frecuencia cardíaca observados en caballos fina sangre chilenos previo y posterior a la competencia de rodeo (*p < 0.05 vs reposo).
Mean values of heart rate in Chilean purebred horses before and after the rodeo competition.
Las concentraciones de los electrólitos Na, Cl y Ca no presentaron variaciones significativas respecto de los valores de reposo. Aunque la concentración de potasio mostró un ligero aumento respecto del reposo a los 15 minutos postejercicio, este no fue estadísticamente significativo. En cambio a las 8 h postejercicio se observó una disminución de la concentración de K (p < 0.05), cambio que es significativo tanto de los valores de reposo como de aquellos obtenidos a los 15 min y a las 24 h postejercicio.
Valores promedios de variables hematológicas y bioquímicas en caballos chilenos en reposo y posterior a la competencia de rodeo (media ± D.E.)
|
|
|||||||||||||
|
|
|
||||||||||||
|
|
|
|
|
||||||||||
|
|
|
|
|
||||||||||
|
|
|||||||||||||
| Hemoglobina |
(g/dL)
|
12.7
|
|
1.8 |
14.2
|
|
2.5* |
11.3
|
|
1.2+ | 12.2 |
|
1.1+ |
| Hematocrito |
(%)
|
37.8
|
|
4.9 |
42.4
|
|
6.1* |
33.8
|
|
3.5+ | 36.8 |
|
3.8+ |
| Proteínas totales |
(g/L)
|
71.4
|
|
6.3 |
76.1
|
|
9.2* |
70.5
|
|
6.2+ | 70.1 |
|
5.2+ |
| Glucosa |
(mmol/L)
|
5.1
|
|
0.7 |
5.7
|
|
1.2* |
5.2
|
|
1.3 | 5.3 |
|
0.6 |
| Acido láctico |
(mmol/L)
|
2.68
|
|
0.92 |
18.7
|
|
12.9* |
3.3
|
|
2.5+ | 2.9 |
|
1.03+ |
| Triglicéridos |
(mmol/L)
|
0.28
|
|
0.16 |
0.41
|
|
0.25* |
0.17
|
|
0.07+ | 0.32 |
|
0.18 |
| Insulina |
(µU/L)
|
8.31
|
|
36.2 |
17.9
|
|
10.7* |
14.7
|
|
16.2 | 4.7 |
|
3.7+. |
| Cortisol |
|
148.5
|
|
47.6 |
198.8
|
|
6.9* |
167.1
|
|
52.9 | 187.9 |
|
66.3 |
| C.K. |
(U/L)
|
62.4
|
|
27.1 |
102.5
|
|
77.1* |
85.2
|
|
42.2 | 67.8 |
|
24.5+ |
| A.S.T. |
(U/L)
|
208.8
|
|
89.0 |
234.2
|
|
142.7 |
155.0
|
|
41.4 | 339.8 |
|
146.5*+. |
| L.D.H. |
(U/L)
|
268.2
|
|
77.3 |
346.1
|
|
147.8* |
265.7
|
|
50.2 | 201.4 |
|
67.2+ |
| Na |
(mEq/L)
|
135.1
|
|
5.9 |
138.0
|
|
5.3 |
135.6
|
|
5.9 | 136.0 |
|
6.2 |
| K |
(mEq/L)
|
3.92
|
|
0.67 |
4.16
|
|
1.24 |
3.12
|
|
0.76*+Ñ | 3.57 |
|
0.52 |
| Cl |
(mEq/L)
|
89.4
|
|
7.6 |
86.8
|
|
6.0 |
90.0
|
|
5.8 | 87.1 |
|
6.1 |
| Ca |
(mEq/L)
|
3.1
|
|
0.47 |
3.02
|
|
0.34 |
2.95
|
|
0.35 | 3.11 |
|
0.36 |
|
|
|||||||||||||
| * p 0.05 vs reposo. + p 0.05 vs 15 min postejercicio. . p < 0.05 vs 6 h postejercicio.Ñ p < 0.05 vs 24 h postejercicio. | |||||||||||||
DISCUSION
Los promedios de las medidas hipométricas de los caballos del presente estudio se encuentran dentro de los valores normales descritos para el estándar de la raza criolla chilena. El peso corporal promedio de los caballos de rodeo fue de 392.1 ± 33.8 kg, el cual está acorde con los valores de alzada y perímetro torácico correspondiente a un caballo de talla mediana y bien proporcionado, características que concuerdan con las de un animal que debe realizar trabajo de corta duración y alta intensidad. En general, se puede señalar que estas medidas corresponden a un animal de baja alzada, con una proporción longitud corporal/perímetro torácico equivalente a un tipo racial de características mesolíneas acordes con las de un caballo musculoso, rápido y agil en sus movimientos, con buenas aptitudes de silla (Beltrán, 1954) y condiciones vaqueras que lo hacen apto para la función que debe cumplir en el rodeo (Pinochet, 1980). Un análisis más detallado acerca de las medidas corporales del caballo criollo se describen en un estudio paralelo al presente trabajo realizado por Cabezas y col. (1996).
La carga total sobre el dorso fue de 92.9 ± 10 kg, la que representa el 23.7 ± 3.7% del peso corporal del animal. Con este peso sobre su dorso el caballo recorre en el apiñadero una distancia de 156 m a una velocidad promedio de 6.95 ± 1.17 m/s. Mientras que en la cancha recorre una distancia de 85 m a una velocidad promedio que fluctúa entre los 7.95 ± 1.13 m/s (28.6 km/h) en la primera corrida y los 4.98/0.98 m/s (17.9 km/h) en la tercera corrida, para atajar un novillo de un peso promedio de 318.4 ± 11.0 kg. Si se considera que en general el bovino es un animal sedentario y que tanto desde el punto de vista anatómico como fisiológico el novillo está menos capacitado que el caballo para realizar trabajo muscular de alta intensidad y resistencia (Hoppeler, 1991), es posible pensar que el principal factor responsable de la disminución de la velocidad durante las diferentes corridas está determinada principalmente por el rendimiento y el nivel de agotamiento que experimenta el novillo. El alto nivel de esfuerzo desarrollado por los caballos del presente estudio estuvo condicionado por el peso del jinete y aperos, que en total representan una carga equivalente al 24% del peso corporal del caballo de rodeo, en comparación al caballo de carreras que trabaja con una carga no superior al 10-12% de su peso corporal. Además, las características de la medialuna son diferentes, ya que se trata de una pista circular, con suelo blando de arena, factores que aumentan la resistencia al desplazamiento del caballo en la carrera, los que asociados al arreo del novillo y al desplazamiento lateral del caballo que va a la mano, es decir aquel que corre a parejas con el novillo, guiándolo, determinan que a pesar de la corta duración del ejercicio éste sea de gran intensidad.
Estos resultados permiten pensar que el rodeo representa para el caballo chileno un notable esfuerzo muscular de corta duración pero de alta intensidad, el cual puede ser intermitente y repetitivo dependiendo del número de series en las cuales puede participar y del nivel de rendimiento que logre durante el transcurso de la competencia. En este sentido, en el primer día de competencia cada collera de caballos puede correr un mínimo de tres animales (si clasifica en su primera participación) o un máximo de 9 novillos si no clasifica en ninguna serie.
Los promedios de frecuencia cardíaca de 110 ± 4 lat/min obervados en los primeros minutos posteriores a la corrida del primer novillo indican una rápida recuperación hacia valores de reposo. En este sentido, es importante considerar lo descrito anteriormente, respecto a que las mayores velocidades registradas durante el rodeo se observan durante el arreo del novillo en el apiñadero y en la primera corrida, donde se alcanzan velocidades promedios que fluctúan entre 7 a 8 m/s. Posteriormente, la velocidad de cada corrida disminuye paulatinamente hasta la entrega del novillo, que se realiza al trote. La disminución de la velocidad durante el rodeo permite al caballo recuperar paulatinamente su frecuencia cardíaca.
En caballos trotones se ha demostrado que cuando estos animales traccionan una carga equivalente a 34 kilopondios (340 N), a una velocidad de 7 m/s, el registro de la frecuencia cardíaca durante el ejercicio alcanza promedios superiores a los 200 lat/min (Gottlieb y col., 1989). Resultados similares describen Thornton y col. (1987), en caballos trotones sometidos a un ejercicio estándar en "treadmill" sobre 3.5º de pendiente y con una carga equivalente al 10% del peso corporal, en los cuales observaron que los valores de V200 (es decir, la velocidad que produce en el caballo una frecuencia cardíaca de 200 lat/min), fluctuaron entre 6.5 a 7.5 m/s, las que fueron significativamente inferiores a los valores observados en caballos que realizaron ejercicios sin carga. Por lo tanto, si se asume que los caballos de rodeo corren a una velocidad similar, con una carga promedio de 92 kg, durante la corrida en la medialuna, éstos debieran presentar incrementos iguales o superiores a los descritos en trotones, lo que indica que a pesar de la corta duración del ejercicio éste representa un esfuerzo físico de alta intensidad.
Se ha demostrado que la frecuencia cardíaca constituye un evaluador confiable para estimar el nivel de esfuerzo que realiza un caballo durante el trabajo muscular en competencia, cualquiera sea la naturaleza del ejercicio (Engelhardt, 1977; Evans, 1985). Por ejemplo, existe una relación estrecha entre el aumento de la velocidad y los cambios observados en los valores de frecuencia cardíaca (Evans, 1985), la que puede ser modificada por las condiciones del suelo, el diámetro de la pista y el peso del jinete (Engelhardt, 1977). Sin embargo, por el hecho de que la frecuencia cardíaca registrada en los caballos del presente estudio corresponde a mediciones realizadas en el animal detenido y dada la rápida recuperación de esta variable una vez terminado el ejercicio, se requieren estudios adicionales en los cuales sea posible medir la frecuencia cardíaca en forma continua durante la corrida del animal para tener una visión más exacta acerca del nivel de intensidad que representa el ejercicio de rodeo para el caballo chileno.
Los promedios de FC de 62.0 ± 3.0 lat/min observados a los 15 min postejercicio indican una rápida recuperación y una adecuada adaptación al trabajo que realizaron los caballos en el rodeo. Estos valores son similares a los descritos en caballos de carreras posterior a competencias de polo (Craig y col., 1985) y en carreras de resistencia (Rose y col., 1980). Es conocido que tanto en el hombre como en los animales la recuperación de la frecuencia cardíaca en el postejercicio está determinada principalmente por la intensidad del ejercicio y por el nivel de acondicionamiento físico del individuo (Cardinet y col., 1963; Aitken y col., 1973; Astrand y Rodahl, 1986). Es por ello que se ha utilizado para evaluar el rendimiento y la aptitud física de los caballos en las competencias (Evans, 1985; Engelhardt, 1977). Así por ejemplo, en caballos de carreras se ha demostrado que aquellos que presentan un buen rendimiento competitivo muestran una recuperación más rápida de la curva de frecuencia cardíaca que aquellos de bajo rendimiento (Milne y col., 1976; Martínez y col., 1994).
Aumentos significativos del volumen globular y de la concentración de hemoglobina se observaron a los 15 minutos posteriores a la corrida del primer animal, cambios que estuvieron asociados a incrementos en la concentración de proteínas totales. Sin embargo, el incremento en los valores de proteínas totales sólo representa un 6.6% respecto del valor de reposo, mientras que el volumen globular y la concentración hemo-globina aumentaron en un 12%; esto indica que una proporción significativa de la hemoconcentración observada es atribuible a la movilización de la reserva de eritrocitos desde el bazo, destinada a favorecer la capacidad transportadora de oxígeno de la sangre para satisfacer la mayor demanda metabólica producida por el aumento del trabajo muscular (Persson, 1967). Mientras que el aumento significativo de la concentración de proteínas es el resultado de la salida de líquidos hacia el compartimiento intersticial y el líquido intracelular (Carlson, 1983, 1987).
Debido a que todos los caballos muestreados dentro de los 15 minutos postejercicio presentaron una importante sudoración, se puede pensar que parte del aumento significativo de la concentración de proteínas plasmáticas refleja una pérdida de líquidos y electrólitos como consecuencia de la competencia de rodeo. Sin embargo, los promedios de concentración de Na, Cl y Ca no mostraron variaciones significativas respecto de sus respectivos valores de reposo. Mientras que la concentración de K disminuyó significativamente en el grupo III (8 horas después de la corrida del primer animal).
Se postula que la mantención de los niveles séricos de Na y Cl es un factor importante para mantener la funcionalidad celular del organismo (Lindinger y Ecker, 1995). A pesar de que las pérdidas de iones por el sudor son importantes, su concentración en el líquido extracelular se mantiene a expensas de las reservas intracelulares, principalmente desde las células musculares y de los eritrocitos (Ferrante y col., 1995). Por lo tanto, la movilización de eritrocitos desde el bazo, inducida por el ejercicio, también representa un aporte importante para mantener los niveles sanguíneos de electrólitos (Ferrante y col., 1995).
Disminuciones significativas en la concentración de potasio han sido observadas en caballos de carreras en el postejercicio especialmente en competencias de velocidad (Krzywanek, 1974) y de resistencia (Carlson y Mansmann, 1974; Snow y col., 1982). Estos cambios son atribuidos en parte a la importante pérdida de K a través del sudor, fluido en el cual alcanza concentraciones 10 a 20 veces superiores a la del plasma (Carlson, 1983). No obstante, una fracción significativa de la caída de la concentración de K plasmático observada en el postejercicio se debe a la recaptación que se produce hacia el interior de la célula muscular y de los eritrocitos circulantes, con el fin de restablecer las concentraciones intracelulares de K, las que disminuyen producto de la salida del catión durante el ejercicio (Rose y col., 1983). Se ha establecido que el músculo en trabajo libera K hacia el medio extracelular, principalmente hacia la circulación, cuya magnitud varía con la intensidad y duración del ejercicio (Harris y Snow, 1988,1992). En el ejercicio, el aumento de K extracelular cumple un rol importante como mediador de la vasodilatación arteriolar para facilitar el flujo de sangre hacia los músculos en trabajo. Por lo tanto, cuando existe una depleción de potasio, tanto su liberación como el flujo de sangre hacia los músculos están reducidos (Freestone y col., 1991) y en humanos se ha asociado a una elevación en la actividad de enzimas y a daño muscular (Knochel y Schlein, 1972).
La concentración de ácido láctico aumentó significativamente desde un valor de reposo de 2.68 ± 0.92 mmol/L a promedios de 18.7 ± 12.9 mmol/L a los 15 min postejercicio. Del total de animales del Grupo II (n = 40), sólo 3 (7.5%) presentaron una concentración igual o inferior a 4 mmoles/L, valor establecido como umbral anaeróbico (Persson y Ullberg, 1974). De los 37 caballos restantes 10, es decir el 25%, tuvieron valores entre 4-10 mmoles/L, 11 (27.5%) entre 10-20 mmoles/L y 16 (40%) presentaron valores de concentración de ácido láctico mayores a 20 mmoles/L, con un valor máximo de 49.6 mmol/L. Estos resultados indican que el 92.5% de los caballos que compitieron en los rodeos realizaron trabajo muscular anaeróbico (Persson y Ullberg, 1974).
Existe amplia evidencia experimental que demuestra que el ejercicio de corta duración, maximal o supramaximal, resulta en la producción intramuscular de cantidades significativas de ácido láctico, cuya concentración, tanto en el músculo como en la sangre, se relaciona exponencialmente con el consumo de oxígeno que acompaña al aumento de la actividad muscular generada por el ejercicio. Por lo tanto, la medición de lactatos en el plasma entrega información valiosa para estimar la deuda de oxígeno producida por el aumento de la actividad muscular (Bayly y col., 1987).
En el caballo, al igual que en el atleta humano, la frecuencia y magnitud de la acumulación de lactatos sanguíneos y/o plasmáticos inducidos por el ejercicio son indicadores utilizados frecuentemente para evaluar la tolerancia al ejercicio y estimar el potencial rendimiento competitivo (Persson y col., 1995). También, constituye uno de los parámetros más utilizados para evaluar el grado o nivel de aptitud física y para optimizar el nivel de entrenamiento, donde aquellos mejor entrenados o más atléticos tienen niveles más bajos de ácido láctico luego de completar un ejercicio estándar (Bayly y col., 1987).
La amplia variación en los valores de concentración de ácido láctico observada en los caballos del presente estudio, como respuesta al trabajo de rodeo, demuestra que éstos presentan diferentes grados de aptitud fisica para realizar trabajo de alta intensidad, lo cual puede ser debido a que no existe un método estándar para lograr una mejor capacidad atlética y competitiva que les permita tolerar en mejor forma la competencia.
Una relación exponencial entre la velocidad desarrollada durante el ejercicio y la concentración de ácido láctico ha sido claramente establecida para el caballo de carreras, en el que se ha observado que a velocidades no mayores de 5.5 a 5.8 m/s, donde la frecuencia cardíaca alcanza valores de 150 lat/min, la concentración de ácido láctico se mantiene en niveles similares a los de reposo. Sin embargo, si la velocidad de carrera se incrementa por sobre estos valores, la concentración de ácido láctico se incrementa exponencialmente y puede alcanzar valores 20 a 50 veces superiores a los de reposo (Engelhardt, 1977). En cambio, en ejercicios de fuerza y resistencia, como es la tracción de un arado de vertedera en faenas de aradura, donde caballos de tiro desarrollan una fuerza equivalente al 17.7% de su peso corporal por períodos de 6 horas diarias, la concentración de ácido láctico se mantiene en valores cercanos a los 4 mmoles/L (Pérez y col., 1996). Por lo tanto, la combinación de carga y velocidad son los factores determinantes en el nivel de esfuerzo físico y en la magnitud de la respuesta fisiológica y metabólica del caballo chileno a la competencia de rodeo. En este sentido, si consideramos que el peso de montura y aperos se mantienen dentro de un rango estándar de 13-14 kg, el peso del jinete puede constituir el principal factor de variación y ser el determinante del rendimiento físico y competitivo del caballo.
El incremento en la concentración de glucosa observado en los caballos de rodeo parece ser una respuesta característica de los atletas humanos (Astrand y Rodahl, 1986) y del caballo (Rose y col., 1980) al ejercicio de alta intensidad y corta duración, la cual está destinada a proveer los requerimientos metabólicos del músculo en trabajo. Es sabido que la concentración de glucosa sanguínea observada durante el ejercicio refleja el balance entre la captación por el músculo en trabajo y su liberación desde el hígado por glucogenolisis (Deldar y col., 1982).
Asociados a las variaciones en la concentración de glucosa se observaron incrementos significativos en la concentración de triglicéridos sanguíneos, resultados que son similares a los descritos por Lucke y Hall (1978) y Pöso y col. (1989) en caballos de carreras sometidos a ejercicios de diferente intensidad. La lipólisis en el tejido adiposo libera ácidos grasos libres y glicerol hacia la circulación. Los ácidos grasos libres son incorporados al músculo para constituir sustratos metabólicos destinados a suplir la demanda de energía generada por la actividad muscular. Debido a que el glicerol no es utilizado durante el ejercicio, la medición de sus niveles plasmáticos provee un índice del grado de lipólisis y del nivel de utilización de ácidos grasos libres por el músculo, que se produce durante el ejercicio (Snow y McKenzie, 1977).
La actividad plasmática de CK, LDH y AST aumentó significativamente sobre los valores de reposo en los caballos sometidos a la competencia de rodeo. De acuerdo a las variaciones observadas, estos aumentos se evidenciaron a los 15 min postejercicio para los promedios de actividad de CK y LDH. En cambio, AST presentó incrementos significativos a las 24 horas postejercicio. El incremento de la actividad de enzimas musculares es una respuesta comÏn observada en el caballo, cualquiera sea el ejercicio al cual es sometido. Así por ejemplo, incrementos significativos han sido observados posterior a carreras de resistencia (Lucke y Hall, 1978; Rose y col., 1980), competencias de polo (Craig y col., 1985) y de salto (Lekeux y col., 1991). Resultados similares han sido observados también en caballos de tiro sometidos a ejercicios de tracción de diferente intensidad y duración (Pérez y col., 1992, 1996). En el caso de CK y LDH por tratarse de enzimas citoplasmáticas de tamaño pequeño y bajo peso molecular, ellas son liberadas hacia la sangre producto del aumento de la permeabilidad de la membrana celular a consecuencia de la hipoxia celular generada por el trabajo muscular anaeróbico (Milne, 1982).
Debido a que los incrementos observados en la actividad de estas enzimas fueron moderados y sus valores se mantienen dentro de rangos normales para la especie, estos resultados parecen indicar que dicho aumento es una consecuencia fisiológica producto de la intensidad del ejercicio. Además, la rápida recuperación de la actividad a valores de reposo en la tarde del primer día de ejercicio parece indicar que su aumento de actividad en el plasma resulta de un cambio de permeabilidad de fibras intactas más que una alteración permanente en la integridad celular (Snow y col., 1982). Sin embargo, no se puede descartar completamente la posibilidad de que en algunos caballos el aumento de la actividad de las enzimas está asociado a daño celular como consecuencia de un mayor esfuerzo muscular. Esto podría explicar el aumento en la actividad de AST, la que por tratarse de una enzima mitocondrial y citosólica su presencia en el plasma es más tardía que CK y LDH, ya que su salida hacia la circulación general requiere de la presencia de daño celular (Kaneko, 1989).
La determinación simultánea de CK y AST en el caballo presenta un valioso potencial diagnóstico y como ayuda de pronóstico de acuerdo a las diferencias en sus tasas de desaparición de su actividad plasmática o sérica. Así por ejemplo, cuando la AST está elevada producto de mionecrosis y la actividad de CK está normal o disminuye indica que la destrucción de células musculares no está activa ((Kaneko, 1989); Harris y col., 1991). El análisis de los valores individuales de AST, observados a las 24 horas postejercicio, demostró niveles de actividad que fluctuaron en rangos de 136 a 693 Ul/L. De los 19 caballos del grupo IV, 10 presentaron valores superiores a 300 Ul/L, establecido por Harris y col. (1991) como el límite superior de valor normal de actividad. Sin embargo, por el incremento moderado observado en los caballos del presente estudio no es posible diferenciar si éstos son de carácter fisiológico o fisiopatológico y, probablemente, más bien están asociados a la falta de entrenamiento o aptitud física de los caballos muestreados. Se ha demostrado que la adaptación que ocurre con el entrenamiento produce una menor liberación de enzimas producto de la reducción de la permeabilidad de la membrana de la célula muscular (Snow y Harris, 1988). Según Harris y col. (1991), para que la actividad plasmática o sérica de enzimas musculares tenga una significancia patológica deben presentar un nivel de actividad mucho mayor que el esperado para la naturaleza del ejercicio y el nivel de aptitud física del caballo.
La respuesta endocrina a la competencia de rodeo de los caballos del presente estudio se caracterizó por presentar incrementos significativos en las concentraciones plasmáticas de cortisol e insulina. El aumento de la concentración de cortisol es una respuesta común del caballo al trabajo o ejercicio, cualquiera sea la actividad que realice (García y Beech, 1986; Church y col., 1987; Wilson y col., 1991). La magnitud de esta respuesta parece reflejar tanto la intensidad como la duración del trabajo realizado (Linden y col., 1991; Wickler y Troy, 1991). Los cambios en la concentración de cortisol observados en el ejercicio parecen estar asociados estrechamente con la variaciones observadas en la concentración de otras hormonas tales como insulina, glucagón, catecolaminas y hormonas tiroideas, las cuales están involucradas en la movilización de sustratos metabólicos en respuesta al mayor trabajo muscular generado por el ejercicio (Thornton, 1985; García y Beech, 1986; Church y col., 1987; Wilson y col., 1991).
El aumento de la concentración de insulina observado a los 15 min postejercicio de rodeo concuerda con los resultados observados en caballos de carreras descritos por Church y col., 1987, quienes han demostrado que a pesar de que la respuesta característica del caballo al ejercicio intenso es una disminución en la concentración de insulina, estos cambios correponden principalmente a aquellos observados durante o inmediatamente después del ejercicio (Dybdall y col., 1980; García y Beech, 1986). En cambio, en el postejercicio se observa una hiperinsulinemia de rebote, en la cual las concentraciones de hormonas aumentan al doble dentro de los 30 min postejercicio y posteriormente declinan paulatinamente hasta lograr concentraciones de reposo (García y Beech, 1986; Church y col., 1987. En el hombre, el ejercicio intenso también condiciona un aumento prolongado en la concentracción de insulina (Wirth y col., 1981). Esta hiperinsulinemia ha sido atribuida a la supresión del efecto inhibitorio que ejerce el sistema simpático sobre las células beta del páncreas (Church y col., 1987. También se ha demostrado que el incremento postejercicio en los niveles plasmáticos de insulina es menor en los caballos entrenados que en los no entrenados Church y col., 1987 Por lo tanto, los cambios en la concentración de insulina parecen ser un indicador del nivel de entrenamiento y un estimador indirecto de los cambios producidos por el ejercicio sobre la actividad del sistema nervioso simpático.
En resumen, se puede señalar que el caballo F.S. chileno se caracteriza por ser un animal de baja alzada, con una relación longitud corporal/perímetro torácico equivalente a un tipo racial de proporciones mesolíneas, con un peso promedio de 392 ± 33 kg, condiciones que definen a un animal adaptado a realizar trabajo de velocidad y fuerza como es el rodeo.
El esfuerzo físico principal que desarrolla durante el rodeo es el de arrear y atajar un novillo de 320 kg de peso, transportando sobre su dorso un peso equivalente al 24% de su peso corporal a velocidades que fluctúan entre los 5 a 8 m/s sobre una pista blanda y circular que determinan modificaciones importantes en la actividad cardiovascular, metabólica y endocrina que corresponden a adaptaciones observadas en caballos sometidos a ejercicios de alta intensidad.
RESUMEN
Se realizó un estudio con el objetivo de caracterizar el nivel de esfuerzo físico y la adaptación fisiológica del caballo fina sangre chileno a la competencia de rodeo. Para el estudio se determinaron las medidas hipométricas, se pesaron los jinetes y aperos. Se midió la medialuna en sus diferentes segmentos y se cronometró el tiempo de carrera con el fin de determinar la velocidad de trabajo. Previo y posterior a la primera corrida de cada caballo, se registró la frecuencia cardíaca y se extrajeron muestras de sangre para la determinación del volumen globular y medir las concentraciones de hemoglobina, proteínas totales, glucosa, ácido láctico, triglicéridos, insulina, cortisol y electrólitos (Na, K, Cl y Ca). También se determinó la actividad de enzimas creatin- kinasa (CK), lactato deshidrogenasa (LDH) y aspartato aminotransferasa (AST).
Los resultados permiten concluir que el caballo F.S. chileno se caracteriza por ser un animal de baja alzada, con una relación longitud corporal/perímetro torácico equivalente a un tipo racial de proporciones mesolíneas, con un peso promedio de 392 ± 33 kg, características que concuerdan con las establecidas para el estándar de la raza criolla chilena.
El esfuerzo físico principal que desarrolla durante el rodeo es el de arrer y atajar un novillo de 318 ± 11 kg de peso, transportando sobre su dorso un peso equivalente al 24% de su peso corporal a velocidades que fluctúan entre los 5 a 8 m/s sobre una pista blanda y circular, que determinan aumentos significativos de la frecuencia cardíaca, en el volumen globular y en las concentraciones de hemoglobina, glucosa, ácido láctico, triglicéridos, insulina y cortisol. Además, de aumentos significativos en la actividad plasmática de CK, LDH y AST. Los cambios más importantes en el balance hidrosalino fueron un aumento en la concentración de proteínas séricas y una disminución en la concentración de K (8 h. postejercicio). Se concluye que el rodeo constituye un ejercicio de fuerza, velocidad y resistencia, que determina importantes modificaciones en la actividad cardiovascular, metabólica y endocrina, que corresponden a adaptaciones observadas en caballos sometidos a ejercicios de alta intensidad.
AGRADECIMIENTOS
Los autores desean agradecer la valiosa colaboración de los señores
presidentes de las Asociaciones de Rodeo de Chillán, Pinto, San Ignacio
y Parral, por las facilidades otorgadas para realizar este estudio dentro de
los recintos de las medialunas respectivas. También se agradece la valiosa
colaboración del Sr. Presidente de la Asociación Chilena de Huasos
de San Carlos.
______________________
Aceptado: 04.03.97.
*Trabajo financiado por Proyecto FONDECYT 1950854
BIBLIOGRAFIA
AITKEN, M.M., J. SANFORD, G. MCKENZIE. 1973. Factors influencing deceleration of heart and respiratory rates after exercise in the horse, Equine Vet. J. 5: 8-14.
ASTRAND, P.O., K. RODAHL. 1986. Fisiología del trabajo físico. Bases fisiológicas del ejercicio. Editorial Médica Panamericana, Bs. Aires.
BAYLY, W.M., B.D. GRANT, R.C. PEARSON. 1987. Lactate concentrations in thoroughbred horses following maximal exercise under field conditions. En: Equine exercise physiology 2. J.R. GILLESPIE and N.E. ROBINSON (eds.), ICEEP Publications. Davis, CA., pp. 426-437.
BELTRAN, J.M. 1954. Ganado caballar. Salvat Editores, S.A., Barcelona.
CABEZAS, I., M. GARCIA, R. GUZMAN, V. MERINO, R. PEREZ. 1996. Características hipométricas, peso corporal y capacidad de carga del caballo Fina Sangre Chileno en competencia de rodeo. Avances en Ciencias Veterinarias. (En evaluación).
CARDINET, G.H., H.E. FOWLER, W.S. TYLER. 1963. Heart and respiratory rates for evaluating performance in horses during endurance trail ride competition, J. Am. Vet. Med. Ass. 143: 1303-1309.
CARDINET, G.H. 1989. Skeletal muscle function. En: Kaneko, J.J. Clinical Biochemistry of domestic animals. 4th edition, Academic Press Inc., San Diego.
CARLSON, G.P., R.A. MANSMANN. 1974. Serum electrolyte and plasma protein alterations in horses used in endurance ride, J. Am. Vet. Ass. 165: 262-264.
CARLSON, G.P. 1983. Thermoregulation and fluid balance in the exercising horse. En: Equine Exercise Physiology. D.H. SNOW, S.G.B. PERSSON & R.J. ROSE (eds.), Granta Editions, Cambridge, pp. 291-309.
CARLSON, G.P. 1987. Blood chemistry, body fluids and hematology. En: Equine Exercise Physiology 2. J.R. GILLESPIE and N.E. ROBINSON (eds.), ICEEP Publications. Davis, CA., pp. 393-425.
CHURCH, D.B., D.L. EVANS, D.R. LEWIS, R.J. ROSE. 1987. The effect of exercise on plasma adrenocorticotrophin, cortisol and insulin in the horse and adaptation with training. En: Equine Exercise Physiology 2. J.R. GILLESPIE and N.E. ROBINSON (eds.) ICEEP Publications, Davis, CA., pp. 506-515.
CRAIG, L., H.F. HINTZ, L.V. SODERHOLM, K.L. SHAW, H.F. SCHRYVER. 1985. Changes in blood constituents accompanying exercise in polo horses, Cornell Vet. 75: 297-302.
DELDAR, A., F.G. FREGIN, J.C. BLOOM, Z. DAVANIPOUR. 1982. Changes in selected biochemical constituents of blood collected from horses participating in a 50-mile endurance ride, Am. J. Vet. Res. 43: 2239-2243.
DYBDALL, N.O., D. GRIBBLE, J.E. MADIGAN, G.H. STABENFEDLT. 1980. Alterations in plasma corticosteroids, insulin and selected metabolites in horses used in endurance ride, Equine Vet. J. 12: 137-140.
ENGELHARDT, W.V. 1977. Cardiovascular effects of exercise and training in horses, Adv. Vet. Sc. & Comp. Med. 21:173-205.
ESMINGER, M.E. 1973. Producción equina. Centro Regional de ayuda técnica. Agencia para el Desarrollo Internacional, Buenos Aires.
EVANS, D.L. 1985. Cardiovascular adaptations to exercise and training, Vet. Clin. North Am., Equine Practice 1: 513-527.
EVANS, D.L., R.C. HARRIS, D.H. SNOW. 1990. Correlation of racing performance with blood lactate and heart rate affer exercise in thoroughbred horses, Equine Vet. J. 25: 441-445.
FERRANTE, P.L., L.E. TAYLOR, J.A. WILSON, D.S. KRONFELD. 1995. Plasma and erythrocyte ion concentrations during exercise in Arabian horses, Equine Vet. J. Suppl. 18: 306-309.
FREESTONE, J.F., K. GOSSET, G.P. CARLSON, G. CHURCH. 1991. Exercise induced alterations in the serum muscle enzymes, erythrocyte, potassium and plasma constituents following feed withdrawal or furosemide and sodium bicarbonate administration in horses, J. Vet. Int. Med. 5: 40-46.
GARCIA, M.C., J.H. BEECH. 1986. Endocrinologic, hematologic and heart rate changes in swiming horses, Am. J. Vet. Res. 47: 2001-2006.
GOTTLIEB, M., B. ESSEN-GUSTAVSSON, H. SKOGLUND-WALLBER. 1989. Blood and muscle metabolic responses to draught work of varying intensity and duration in horses, Res Vet. Sci. 47: 102-109.
HARRIS, P., D.H. SNOW. 1988. The effects of high intensity exercise on the plasma concentration of lactate, potassium and other electrolytes, Equine Vet. J. 20: 109-113.
HARRIS, P.A., D.H. SNOW, T.R. GREET, P.D. ROSSDALE. 1991. Some factors influencing plasma AST/CK activities in thoroughbred racehorse, Equine Vet. J. Suppl. 9: 66-71.
HARRIS, P., D.H. SNOW. 1992. Plasma potassium and lactate concentrations in thoroughbred horses during exercise of varying intensity, Equine Vet. J. 23: 220-225
HOPPELER, H. 1991. What makes horses superior athletes? En: Proceedings of the International Conference on Equine Sport Medicine. Ed. P. Kallings, Dept. Medicine & Surgery, Faculty of Veterinary Medicine, Swedish University of Agricultural Sciences, Uppsala, pp. 7-12.
KANEKO, J.J. 1989. Clinical biochemistry of domestic animals. 4th ed., Academic Press, San Diego, pp. 480-486.
KNOCHEL, J.P., E.M. SCHLEIN. 1972. On the mechanism of rhabdomyolysis in potassium depletion, J. Clin. Invest. 51: 1750-1758.
KRZYWANEK, H. 1974. Lactic acid concentrations and pH values in trotters after racing, J. S. Afr. Vet. Ass. 45: 355-360.
KRZYWANEK, H., D.W. MILNE, A.A. GABEL, G.L. SMITH. 1976. Acid-base values of standardbred horses recovering from strenous exercise, Am. J. Vet. Res. 37: 291-294.
LEKEUX, P., T. ART, A. LINDEN, D. DESMECHT, H. AMORY. 1991. Heart rate, hematological and serum biochemical responses to show jumping, Equine Exercise Physiology 3: 385-390.
LINDEN, A., T. ART, H. AMORY, D. DESMECHT, P. LEKEUX. 1991. Effect of 5 different types of exercise, transportation and ACTH administration on plasma cortisol concentration in sport horses, Equine Exercise Physiology, 3: 391-396.
LINDINGER, M.I., G.L. ECKER. 1995. Ion and water losses from body fluids during a 163 km endurance ride, Equine Vet. J. Suppl. 18: 314-322.
LUCKE, J.N., G.M. HALL. 1978. Biochemical changes in horses during a 50-mile endurance ride, Vet. Rec. 102: 356-358.
MARTINEZ, R., E. GONZALEZ, B. CAMPOS. 1987. Lactacidemia post-trabajo supramaximal en equinos F.S.C. de alto rendimiento, Arch. Med. Vet. 19: 57-62.
MARTINEZ, R. 1989. Bases fisiológicas para el manejo hípico del equino F.S.C., MonografÕas Med. Vet. 11: 20-41.
MARTINEZ, R., G. FILGUEIRA, R. CARRILLO, A. WHITE. 1994. Registro continuo de frecuencia cardíaca en equinos de diferente aptitud competitiva sometidos a ejercicio, Monografías Med. Vet. 16: 81-86.
MILNE, D.W. 1982. Biochemical parameters for assessment of conditioning in the horse, Proc. Am. Assoc. Equine Pract. 28: 49-53.
MILNE, D.W., R.T. SKARDA, A.A. GABEL, L.G. SMITH, K. AULT. 1976. Effect of training on biochemical values in standardbred horses, Am. J. Vet. Res. 37: 285-290.
PEREZ, R., S. RECABARREN, P. VALDES, E. HETZ. 1992. Biochemical and physiological parameters and estimated work output in draught horses pulling loads for long periods, Vet. Res. Comunic. 16: 231-246.
PEREZ, R., I. CABEZAS, R. GUZMAN, M. GARCIA, C. CHAVARRIA, R. SOTO. 1993. Comparación de características hipométricas de caballos mestizos descendientes de potros fina sangre y criollos de tiro, Av. Cienc. Vet. 8: 119-128.
PEREZ, R., S. VALENZUELA, V. MERINO, I. CABEZAS, M. GARCIA, R. BOU, P. ORTIZ. 1996. Energetic requirements and physiological adaptation of draught houses to ploughing work, Animal Science 6(2): 343-351.
PERSSON, S.G.B. 1967. On blood volume and working capacity in horses. Studies of methodology and physiological and pathological variations, Acta Vet. Scand. Suppl. 19: 1-189.
PERSSON, S.G.B., L.E. ULLBERG. 1974. Blood volume in relation to exercise tolerance in trotters, J.S. Afr. Vet. Ass. 45: 293-299.
PERSSON, S.G.B., B. ESSEN-GUSTAVSSON, P. FUNKQUIST, L. BLANCO-ROMERO. 1995. Plasma, red cell and whole blood lactate concentrations, during prolonged treadmill exercise at VLA4, Equine Vet. J. Suppl. 18:104-107.
PINOCHET, J.L. 1980. Estudio hipométrico y morfológico del caballo de raza criolla chilena y su posible cambio tipológico. Tesis. Universidad de Chile. Facultad de Ciencias Veterinarias, Santiago, Chile.
PÖSO, A., E. VILJANEN-TARIFA, T. SOVERI, H. OKSANEN. 1989. Exercise-induced transient hyperlipidemia in the racehorse, J. Vet. Med. Serie A. 36: 603-611.
ROSE, R.J., R. PURDUE, W. HENSLEY. 1977. Plasma biochemistry alterations in horses during an endurance ride, Equine Vet. J. 9: 122-126.
ROSE, R.J., J.E. ILKIW, K.S. ARNOLD. 1980. Plasma biochemistry in the horse during 3-day event competition, Equine Vet. J. 12: 132-136.
ROSE, R.J., D.R. HODGSON, D. SAMPSON, W. CHAN. 1983. Changes in plasma biochemistry in horses competing in a 160 km endurance ride, Aust. Vet. J. 60: 101-105.
ROSE, R.J. 1986. Endurance exercise in the horse. A review. Part I and II, Br. Vet. J. 142: 532-552.
ROSE, R.J., D.R. HODGSON. 1993. Manual of equine Practice. W.B. Saunders Co., Philadelphia, Pennsylvania, 486 pp.
SNOW, D.H., G. MACKENZIE. 1977. Some metabolic effects of maximal exercise in the horse and adaptations with training, Equine Vet. J. 9: 134-140.
SNOW, D.H., M.G. KERR, M.A. NIMMO, E.M. ABBOTT. 1982. Alterations in blood, sweat, urine and muscle composition during prolonged exercise in the horse, Vet. Rec. 110: 377-384.
SNOW, D.H., P. HARRIS. 1988. Enzymes markers for the evaluation of physical fitness and training of racing horses. En: GOLDBERG, D.M., D.W. MOSS, E. SCHMIDT and F.W. SCHMIDT. Enzymes-Tools and target. Advances in Clinical Enzymology. Basel, Karger, Vol. 6, pp. 251-258.
THORNTON, J.R. 1985. Hormonal response to exercise and training. Vet. Clin. N. Am. Equine Pract. 1: 477-496.
THORNTON, J.R., J. PAGAN, S. PERSSON. 1987. The oxygen cost of weight loading and inclined treadmill excercise in the horse. En: Equine exercise physiology 2. J.R. GILLESPIE y N.E. ROBINSON (eds.). ICEEP Publications, Davis, CA., pp. 206-215.
VALDIVIA, V.H. 1995. Estudio hipométrico y conformacional del caballo fina sangre chileno de la cuarta región. Memoria de título, Facultad de Medicina Veterinaria, Universidad de Concepción, Chillán, 85 pp.
WICKLER, S.J., W. TROY. 1991. Blood volume, lactate and cortisol in exercising Arabian equitation horses, Equine Excercise Physiology 3: 397-401.
WILSON, W.D., S. KINGERY, D.H. SNOW. 1991. The effect of training on adrenocortical function in thoroughbred racehorses, Equine Exercise Physiology 3: 482-489.
WIRTH, A., C. DIEHM, H. MAYER, H. MORL, I. VOGEL, P. BJORNSTORP, G. SCHLIERF. 1981. Plasma c-peptide and insulin in trained and untrained subjects, J. Appl. Physiol. 50: 71-77.