R. PEREZ, M.V., M.Sc.; C. BREVIS, M.V.; M. GARCIA, M.V.; V.
MERINO, B.Q., M.Sc.;
S. VALENZUELA, B.Q.; R. GUZMAN, M.V.; A. ISLAS, M.V., M.Sc.; I. CABEZAS, M.V.
Departamento de Ciencias Clínicas, Facultad de Medicina
Veterinaria, Universidad de Concepción,
Casilla 537, Campus Chillán, Chile
SUMMARY
Fibre composition and capillary supply of the Gluteus medius
muscle in
resting Chilean criollo horses
A study to analyze fibre composition and capillary supply of the Gluteus medius muscle of the Chilean criollo horse under resting conditions was carried out. Six adult (5-10 year old), clinically healthy Chilean criollo horses were sampled through a muscle biopsy technique during the period previous to the rodeo competition. Muscular biopsies were obtained from the left Gluteus medius at 6 cm of depth below the fascia. Muscle samples were serially sectioned and stained for myofibrilar ATP-ase and amylase-PAS, in order to identify types I, IIA and IIB fibres and to evaluate the fibre type distribution, the cross sectional area and capillary density.
The results showed that the Chilean criollo horse has a greater proportion of fast twitch type II fibres than type I, whereas type IIB fibres (42.5%) showed a slightly greater proportion than type IIA (39.0%), which suggests that this breed of horses is best adapted to muscular work of speed and resistance. The greatest frequency of type IIB fibres and the low capillary density (253 ± 26 capillaries/mm2) seem to demonstrate that these characteristics are associated to a loss of fitness due to a prolongued resting period to which the horses were subjected.
Palabras claves: fibra muscular, densidad capilar, caballos, histoquímica.
Key words: muscle fibre, capillary supply, horses, histochemistry.
INTRODUCCION
El músculo esquelético está constituido por fibras musculares individuales, las cuaIes poseen propiedades contráctiles y metabólicas diferentes (Valberg y Essén-Gustavsson, 1987). Sin embargo, las fibras que pertenecen a una misma unidad motora, es decir, aquellas que son inervadas por las ramas de una misma motoneurona, tienen propiedades idénticas (Snow, 1983).
Mediante la técnica de biopsia muscular ha sido posible caracterizar la respuesta metabólica del músculo esquelético al ejercicio y se ha demostrado que ésta es dependiente tanto del tipo de fibra, la proporción de los diferentes tipos de fibra dentro del músculo, como de la capacidad oxidativa y glicolítica de ellas (Ronéus y col., 1991).
El Gluteus medius ha sido el músculo más frecuentemente examinado por los fisiólogos para estudiar el efecto del ejercicio en el atleta equino, debido a que se caracteriza por ser un músculo de gran volumen, fácil acceso y principalmente por el hecho de que se trata de un músculo que participa en la propulsión del miembro posterior durante el trote y el galope (Lindholm y Piehl, 1974; López-Rivero y col., 1992).
Las características metabólicas de las fibras musculares pueden ser determinadas mediante el análisis de indicadores de su capacidad glicolítica y oxidativa. Además, su actividad contráctil puede ser determinada por la medición de la actividad de la miosina ATP-asa miofibrilar (Essén-Gustavsson, 1991). Sobre la base de la actividad de la miosina ATP-asa a pH 9.4 se distinguen dos tipos de fibras, las denominadas fibras tipo I o fibras de contracción lenta o baja actividad ATP-asa y las fibras tipo II que son de contracción rápida y alta actividad (López, 1993). Además, se ha demostrado que en la mayoría de las especies estudiadas, estos dos tipos de fibras están relacionadas con la contractibilidad del músculo, siendo las fibras de tipo I las que presentan tiempos de contracción y relajación más lentos que las fibras tipo II, además de ser más resistentes a la fatiga (Snow, 1983). A su vez las fibras tipo II han sido subclasificadas en los tipos IIA, IIB y IIC de acuerdo a la labilidad de su actividad miosina ATP-asa luego de la incubación a pH ácido en rangos de 4 a 5 (Brooke y Kaiser, 1970).
Debido a su alta capacidad oxidativa las fibras tipo I son reclutadas en los ejercicios submaximales prolongados, teniendo la capacidad de entregar energía de manera continua al degradar glicógeno y triglicéridos. En cambio las fibras tipo IIA y IIB son reclutadas en ejercicios de alta intensidad y corta duración debido a su mayor capacidad glicolítica, que le permite obtener energía más rápidamente. A su vez, en las fibras tipo IIB, que tienen menor capacidad oxidativa, la energía es proporcionada por la degradación de glicógeno con la consiguiente acumulación de lactato (Lindholm y Piehl, 1974; Ruz-Fernández, 1993).
Se ha demostrado que las características de los diversos tipos de fibras musculares varían entre razas equinas (López-Rivero y col., 1989; Snow y Guy, 1980). El caballo Cuarto de Milla tiene alta proporción de fibras de contracción rápida, aproximadamente 100%, los Fina Sangre de Carrera (FSC) tienen menor proporción de fibras tipo II y de menor tamaño que los de raza estándar Snow y Guy, 1980). El caballo Arabe tiene mayor porcentaje de fibras tipo I y IIA, de tamaño pequeño y homogéneo, excelentes atributos para el ejercicio de resistencia (Ronéus, 1992). El caballo Andaluz posee un porcentaje de fibras tipo I intermedio entre las 2 razas anteriores, una mayor proporción de fibras tipo IIB no oxidativas y fibras musculares de mayor dimensión (López-Rivero y col., 1990). Sin embargo, no existen antecedentes que describan las características y distribución de los diferentes tipos de fibras presentes en los músculos del caballo chileno.
Mediante técnicas histoquímicas es posible investigar el tipo de fibra, el área de las fibras, la densidad capilar, la actividad enzimática y la concentración de sustratos intramusculares presentes en un músculo determinado (Essén-Gustavsson y col., 1989). En consideración a que no existen antecedentes que describan las características de las fibras de los músculos del caballo chileno, se realizó el presente estudio con el objetivo de estudiar la composición miofibrilar y la densidad capilar del músculo Gluteus medius en condiciones de reposo.
MATERIAL Y METODOS
Animales. Se utilizaron seis caballos criollos chilenos finos inscritos, adultos, 4 machos y 2 hembras, entre 5-10 años de edad, los que fueron muestreados en condiciones de reposo, luego de un período prolongado de descanso (2 a 6 meses) sin desarrollar actividad física, previo al inicio de la temporada de rodeo.
Biopsias musculares. Las biopsias musculares fueron extraídas desde el músculo Gluteus medius izquierdo. Estas fueron obtenidas con una aguja de 6 mm de diámetro interno según la técnica descrita por Lindholm y Piehl (1974). El sitio exacto se determinó buscando el punto medio de una línea recta trazada entre el punto más alto de la tuberosidad coxal y la base de la cola. Se depiló y desinfectó un área de 3 cm2 de piel, ubicada 10 cm dorsocaudal al trocanter major del tuber coxal, la que fue anestesiada con 2-3 ml de Lidocaína (Dimecaína al 2%, Instituto Bioquímico Beta, Santiago). Se realizó una incisión sobre la piel de 10 mm de largo, a través de la cual se introdujo la aguja de biopsia muscular. Cada biopsia fue extraída de una profundidad estándar de 6 cm y todas fueron realizadas por la misma persona. Luego de extraídas las muestras fueron embebidas en talco, para evitar artefactos de congelación (Valberg y col., 1989), posteriormente fueron congeladas en nitrógeno líquido y mantenidas a -180° C hasta su análisis. Posterior a los procedimientos de recolección de biopsias los caballos fueron sometidos a una inspección visual con el fin de determinar algún grado de claudicación o alteraciones en el caminar atribuible a los procedimientos de extracción de las muestras.
Análisis histoquímico. De las muestras de músculo extraídas se realizaron cortes transversales seriados de 10 µ de espesor en un criostato a -20° C, los que posteriormente fueron incubados por 5 minutos a temperatura ambiente para demostrar la actividad miosina adenosin trifosftasa (mATPasa) miofibrilar a pH 9.4 tras preincubación ácida en buffer acetato (pH 4.3 y 4.6) y alcalina en buffer glicina (pH 10.3), según técnica descrita por Anderssen y Henricksson (1977). Según sus patrones de intensidad de reacción enzimática por mATPasa las fibras fueron clasificadas como tipo I (contracción lenta), o tipos IIA, IIB y IIC (contracción rápida). Las fibras tipo IIC fueron consideradas como fibras IIB, dado que éstas fueron escasas o estuvieron ausentes en la mayoría de las biopsias (Rivero y col., 1993a). Un área representativa de cada biopsia (entre 0.25 y 0.5 mm2 ) conteniendo al menos 200 fibras fue examinada sistemáticamente en los cortes seriados reaccionantes a la mATPasa tras preincubación ácida a pH 4.6 (Karlström y col., 1995).
Otra serie de cortes de 20 µ de espesor fueron teñidas con alfa amilasa-PAS para la determinación del número de capilares por área; para ello se seleccionó un área representativa de cada biopsia conteniendo al menos 200 fibras.
La composición fibrilar, el área de sección transversal de las fibras musculares estriadas y la densidad capilar de las fibras, se determinó mediante un sistema de análisis de imagen computarizado, diseñado para el estudio de fibras musculares esqueléticas (Bio-Rad Scan Beam, Hadsund, Dinamarca), perteneciente a la Facultad de Medicina Veterinaria, Universidad Sueca de Ciencias Agrícolas, Uppsala, Suecia(Karlström y col., 1995). .
Análisis estadístico. La media y la desviación estándar de cada variable fueron obtenidas mediante un programa computacional a través de procedimientos estadísticos convencionales.
RESULTADOS Y DISCUSION
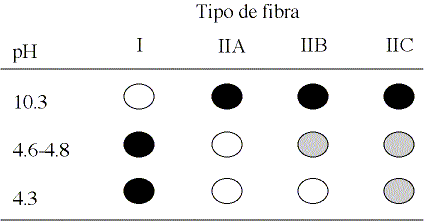
Composición fibrilar. En el cuadro 1 se muestran los promedios y la desviación estándar de la frecuencia de distribución relativa de los diferentes tipos fibrilares que componen el músculo G. medius del caballo chileno, determinados mediante la técnica de la mATPasa miofibrilar. Un esquema con las características tintoriales que muestran las fibras musculares incubadas a 3 valores de pH diferentes se muestra en la figura 1.
Figura 1. Representación esquemática de los diferentes
tipos de fibras musculares determinadas mediante la intensidad de reacción
a mATPasa fibrilar tras preincubación a pH ácido (4.3-4.8) o alcalino
(10.3).
Diagram which shows the different muscle fibre types through the intensity of
reaction to mATPase after acid (pH 4.3-4.8) or alcaline (pH 10.3) preincubation.
Se encontró un 18.5 ± 2.4% de fibras tipo I de contracción
lenta, un 39 ± 6.7% de fibras tipo IIA y un 42.5 ± 6.4% de fibras
tipo IIB. Las fibras de menor diámetro de sección transversal
fueron las tipo I con un promedio de 43.2 ± 7.7 mm seguido de las tipo
IIA con 49.0 ± 6.9 mm, siendo las de mayor diámetro las tipo IIB
con 59.6 ± 7.2 mm. El área de sección transversal ocupada
por cada tipo de fibra fue de 2581 ± 897 mm2 para las fibras
tipo I, 3125 ± 794 mm2 las de tipo IIA y 4789 ± 781
mm2 las de tipo IIB (cuadro 1).
En la figura 2a se muestra una microfotografía de
un corte transversal de una biopsia muscular de Gluteus medius extraída
desde caballos fina sangre chilenos; en ella se aprecia la mayor proporción
de fibras de contracción rápida tipo IIA y IIB.
En el (cuadro 1) se puede observar que del área total estudiada de cada corte seriado de biopsia muscular las fibras tipo I sólo ocupan 12.9 ± 4.6%, mientras que el mayor porcentaje está representado por el área ocupada por las fibras tipo IIB, las cuales representan el 54.6 ± 10.2% del área total, lo que concuerda con el mayor diámetro y la mayor área de sección transversal que presentan estas fibras.
|
Frecuencia relativa, diámetro y área de
sección transversal de las fibras musculares |
|||
|
|
|||
|
Tipo de fibra
|
|||
|
I
|
IIA
|
IIB
|
|
|
|
|||
| Frecuencia relativa (%) |
18.5 ± 2.4
|
39.0 ± 6.7
|
42.5 ± 6.4
|
| Diámetro fibra (mm) |
43.2 ± 7.7
|
49.0 ± 6.9
|
59.6 ± 7.2
|
| Area de fibra (mm2) |
2581 ± 879
|
3125 ± 794
|
4789 ± 781
|
| Area de distribución | |||
| relativa (%) |
12.9 ± 4.6
|
32.5 ± 8.5
|
54.6 ± 10.2
|
|
|
|||
Capilarización. En la figura 2b se muestra
la densidad capilar de las fibras musculares del músculo Gluteus medius
del equino fina sangre chileno. El recuento de densidad capilar determinó
un promedio de 252.6 ± 25.6 capilares/mm2.
No existen reportes previos que describan la composición miofibrilar y la densidad capilar del músculo Gluteus medius del equino fina sangre chileno, por lo tanto los resultados obtenidos deben ser comparados con aquellos descritos para razas de aptitud similar como es el caballo Andaluz (López-Rivero y col., 1989, 1992, 1993; López y col., 1993).
Con el fin de disminuir los factores de variación en los resultados, en el presente trabajo se tuvo cuidado de mantener una profundidad estándar de 6 cm para la extracción de las biopsias. En el caballo Andaluz se ha demostrado que aunque el músculo Gluteus medius puede ser muestreado a una profundidad de 8-10 cm, parece ser que 6 cm es la profundidad más adecuada, ya que en las porciones más profundas este músculo está compuesto casi exclusivamente por fibras tipo I y IIA (López-Rivero y col., 1992).
Por lo tanto, se puede concluir que la profundidad de muestreo utilizada en los caballos del presente estudio es la más adecuada, ya que permite obtener biopsias con una distribución más uniforme de los distintos tipos fibrilares (Rivero y col., 1995).
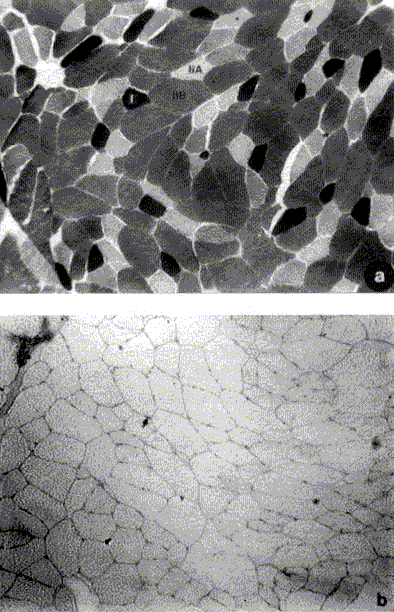
Figura 2. Microfotografías de biopsia muscular de Gluteus
medius de equino fina sangre chileno.
a) Composición fibrilar según reacción
a la miosina ATPasa tras preincubación ácida a pH 4.6 (aumento
100x).
b) Densidad capilar de las fibras musculares evidenciada
mediante la tinción amilasa-PAS (aumento 100x). (La flecha indica un
capilar).
Micrographs of cross-sectional areas of the Gluteus medius muscle in Chilean
horses.
a) Fiber type identification according to mATPase after acid (pH 4.6) preincubation
(x100).
b) Capillary density of Gluteus medius muscle fibres evidenced by amylase-PAS
reaction (x100). (The arrow indicates a capillar).
Los resultados obtenidos sobre la composición miofibrilar del músculo G. medius del caballo chileno indican una proporción promedio de fibras tipo I de contracción lenta cercana al 20% y un 80% de fibras tipo II de contracción rápida. Estos resultados concuerdan con los descritos por Ronéus y col. (1991), quienes describen una proporción similar en estudios realizados en caballos trotones. Esta proporción entre fibras tipo I/tipo II sería característica de aquellas razas que realizan trabajo muscular de velocidad y resistencia (Guy y Snow, 1981). Es decir, el hecho de que los caballos tengan una mayor proporción de fibras de contracción rápida significa que el músculo esquelético está diseñado para realizar trabajo de alta intensidad (Lindholm y Piehl, 1974). El caballo fina sangre de carreras, el cual ha sido seleccionado para realizar trabajo muscular de alta velocidad, presenta entre un 10-15% de fibras de contracción lenta y un 85-90% de contracción rápida (Ronéus, 1992). Por el contrario, en razas pesadas de tiro como el Heavy Hunter la proporción de fibras tipo I aumenta a promedios de 31%, indicando que estas razas están mejor capacitadas para realizar trabajo de fuerza y resistencia a baja velocidad (Snow, 1983).
Algunos autores consideran que la proporción entre fibras tipo I y tipo II está determinada gené-ticamente para las diferentes razas de caballos y que ésta es poco influenciada por el entrenamiento (Snow, 1983); Rivero y col., 1993a y b). El caballo chileno constituye un tipo racial que en su origen proviene de los caballos andaluces traídos por los españoles durante la conquista de América (Seifert, 1971). Sin embargo, los resultados del presente estudio indican que la proporción de fibras tipo I es inferior a la descrita para el caballo Andaluz, que fluctúa entre valores de 27-32% (López-Rivero y col., 1990, 1992) a una profundidad similar a la utilizada en el presente estudio.
Es sabido que las fibras tipo I están bien dotadas tanto desde el punto de vista estructural como funcional y bioquímico para resistir la fatiga, por lo que resultan esenciales en las actividades musculares de larga duración y baja intensidad como son por ejemplo funciones posturales o ejercicios de resistencia (López y col., 1993). En cambio las fibras tipo II son de contracción rápida y vigorosa, más adecuadas para ejercicios de corta duración y alta intensidad (Essén-Gustavsson, 1991).
Las fibras tipo II de contracción rápida son agrupadas en los subtipos IIA y IIB, respectivamente, de acuerdo a sus características metabólicas y capacidad oxidativa (Essén-Gustavsson, 1986). En el presente estudio se encontró una proporción de fibras tipo IIB ligeramente superior a las de tipo IIA, resultado que se explicaría por el hecho de que las muestras fueron extraídas desde animales que se encontraban en un período de reposo prolongado previo al inicio de la temporada de rodeo. Resultados similares han sido descritos en caballos trotones por Essén-Gustavsson y Lindholm (1985), quienes concluyen que en el caballo adulto una baja proporción entre fibras tipo IIA/IIB parece ser una característica frecuentemente observada en biopsias musculares provenientes de animales que han estado inactivos por largos períodos. Por lo tanto, no sólo factores genéticos sino que también factores ambientales como una mayor o menor actividad física pueden ser de importancia para determinar la respuesta adaptativa en el potencial oxidativo y en la composición fibrilar de un determinado músculo esquelético (Essén-Gustavsson, 1986).
Se ha observado que en caballos de carreras se presenta una elevada proporción entre los tipos IIA/IIB y una alta capacidad oxidativa, características que corresponden a animales de gran capacidad atlética (Lindholm y col., 1983).
Las fibras tipo I fueron las que presentaron un menor tamaño, con un diámetro y un área de sección transversal inferiores a las que se observaron en las fibras tipo II. Desde el punto de vista teórico, las fibras de menor tamaño están mejor habilitadas para realizar metabolismo oxidativo que las de mayor tamaño ya que un diámetro pequeño facilita una mejor difusión de oxígeno y nutrientes; además de facilitar la excreción de lactatos y productos metabólicos de desecho cuya acumulación intrafibrilar es causa de fatiga (Essén-Gustavsson y col., 1984).
De las fibras tipo II analizadas en el presente estudio, las tipo IIB fueron las que presentaron un mayor diámetro y una mayor área de sección transversal, características que asociadas a una mayor frecuencia relativa determinan que estas fibras ocupen el 54.6% del área total de distribución relativa de las fibras del músculo G. medius del equino fina sangre chileno.
Desde el punto de vista mecánico, la generación de fuerza muscular está relacionada con el área de sección transversal de las fibras involucradas en la función. Por lo tanto, la mayor área de sección transversal de las fibras tipo IIB, en comparación con las fibras tipo I y IIA, significa que si un número igual de fibras tipo IIB son reclutadas durante el ejercicio ellas producen una mayor fuerza (Valberg y col., 1985).
Aunque desde el punto de vista atlético la actividad física que realiza el caballo chileno en la competencia de rodeo no ha sido aún caracterizada, se asume que se trata de un ejercicio de corta duración y alta intensidad en que se combinan factores de fuerza, velocidad y resistencia. Por lo tanto, se puede especular que durante el transcurso de sus 500 años de evolución en un territorio montañoso y difícil (Leigh, 1983), el caballo criollo experimentó cambios conformacionales que pudieron modificar su composición miofibrilar que difiere de la raza de origen que es el caballo Andaluz. Sin embargo, se requieren estudios comparativos adicionales para comprobar esta hipótesis.
La densidad capilar de las fibras del músculo G. medius del caballo criollo fue de 253 ± 26 capilares/mm2, promedio que es inferior al descrito para el caballo Andaluz que presenta rangos de densidad capilar que fluctúan entre 500-700 capilares/mm2 (Rivero y col., 1995).
La estimación de la densidad capilar depende tanto de la relación fibra-capilar como del tamaño fibrilar (Karlström y col., 1992). Además, la irrigación de cada tipo de fibra está estrechamente relacionada a la capacidad oxidativa de la fibra, así por ejemplo, las fibras tipo I tienen una mayor irrigación que las tipo IIA, las que a su vez están irrigadas por un mayor número de capilares que las IIB (Karlström y col., 1991, 1995). Por lo tanto, aquellos músculos que presentan una gran cantidad de fibras tipo IIB muestran una baja irrigación capilar, tanto por el mayor diámetro fibrilar que ellas presentan como por una menor relación fibra-capilar (Karlström y col., 1991).
La menor densidad capilar observada en los caballos del presente estudio probablemente está asociada al hecho de que en el caballo criollo existe una alta proporción de fibras tipo IIB, las que al poseer una mayor área de sección transversal determinan una menor densidad capilar como ha sido demostrado en caballos trotones (Valberg, 1987; Karlström y col., 1991)..
Se ha demostrado, tanto en el hombre (Anderssen y Henriksson, 1977) como en el caballo (Karlström y col., 1991), que los principales efectos del entrenamiento resultan en una considerable proliferación de la red capilar que irriga las fibras musculares, cambio que está asociado a un incremento en el porcentaje de fibras tipo IIA a expensas de las fibras tipo IIB que disminuyen en frecuencia y tamaño (Andersen y Herinksson, 1977). Por lo tanto, es posible pensar que la baja densidad capilar observada en los caballos del presente estudio esté asociada a la falta de aptitud física debido al reposo prolongado al cual estuvieron sometidos estos animales, que determinó una mayor proporción de las fibras tipo IIB asociada a una baja densidad capilar.
RESUMEN
Se realizó un estudio con el objetivo de analizar la composición fibrilar y la densidad capilar del músculo Gluteus medius del equino fina sangre chileno en reposo. Para el estudio se muestrearon 6 caballos chilenos adultos (5-10 años), durante el período de descanso previo al inicio de la temporada de rodeo.
Biopsias musculares fueron extraídas desde una profundidad estándar de 6 cm. Mediante la técnica de la miosina ATPasa miofibrilar se determinó la distribución de los distintos tipos de fibras musculares, las que según sus patrones de reacción fueron clasificadas en los tipos I, IIA y IIB. Además, la densidad capilar se determinó mediante la técnica de alfa amilasa PAS.
Los resultados muestran una mayor proporción de fibras de contracción
rápida tipo II, dentro de ellas las tipo IIB son más frecuentes
(42.5%) que las tipo IIA (39.0%). Esta distribución en la composición
fibrilar significaría que el músculo G. medius del caballo chileno
está mejor adaptado para realizar trabajo de velocidad y resistencia.
La mayor frecuencia de fibras tipo IIB y una baja densidad capilar (253 ±
26 capilares/mm2) permiten pensar que estas características
están asociadas a una falta de aptitud física producto de un reposo
prolongado.
AGRADECIMIENTOS
Los autores expresan su agradecimiento al Dr. Sune Persson, de la Facultad de Medicina Veterinaria, Universidad Sueca de Ciencias Agrícolas, Uppsala, Suecia, las facilidades otorgadas para realizar el análisis de las fibras musculares. También se agradece la colaboración del Dr. Hernán Urrutia por sus gestiones ante los propietarios de caballos chilenos para facilitar el acceso a los criaderos para la extracción de muestras.
Aceptado: 04.03.97.
Trabajo financiado por Proyecto FONDECYT 1950854.
BIBLIOGRAFIA
ANDERSSEN, P., J. HENRIKSSON. 1977. Capillary supply of the quadriceps femoris muscle of man: Adaptative response to excercise, J. Physiol. 270: 677-690.
BROOKE, N.H., K.K. KAISER. 1970. Muscle fiber types. How many
and what kind?, Arch. Neurol. 23: 369-379.
ESSEN-GUSTAVSSON, B., K. KARLSTRÖM, A. LINDHOLM. 1984.
Fibre types enzime activities and substrates utilization in skeletal muscles
of horses competing in endurance rides, Equine Vet. J. 16: 197-202.
ESSEN-GUSTAVSSON, B., A. LINDHOLM. 1985. Muscle fibre characteristics
of active and inactive standardbred horses, Equine Vet. J. 17: 434-438.
ESSEN-GUSTAVSSON, B. 1986. Activity and inactivity related
muscle adaptation in the animal kingdom. En: SALTIN, B. (ed.). Biochemistry
of exercise VI. Human Kinetic Publishers, Champaigne, IL, pp. 435-444.
ESSEN-GUSTAVSSON, B., D. McMIKEN, K. KARLSTRÖM, A. LINDHOLM,
S. PERSSON, J. THORNTON. 1989. Muscular adaptation of horses during intensive
training and detraining, Equine Vet. J. 21: 27-33.
ESSEN-GUSTAVSSON, B. 1991. Training effect on skeletal muscle.
En P. KALLINGS (ed.) Proceedings of the International Conference on Equine Sport
Medicine. Almquist and Wiksell Tryckeri, Uppsala, pp. 18-23.
GUY, P.S., D.A. SNOW. 1981. Fiber type and enzyme activities
of the gluteus medius in different breed of horse. En: POORTMANS and G. NISET
(eds.) Biochemistry of exercise IV-B. University Park Press, Baltimore, pp.
271-282.
KARLSTRÖM, K., B. ESSEN-GUSTAVSSON, A. LINDHOLM, S.G.B.
PERSSON. 1991. Capillary supply in relation to muscle metabolic profile and
cardiocirculatory parameters. En: S.G.B. PERSSON, A. LINDHOLM, L.B. JEFFCTOT
(eds.). Equine Exercise Physiology 3. ICEEP Publications, Davis, California,
pp. 239-244.
KARLSTRÖM, K., B. ESSEN-GUSTAVSSON, H. HOPPELER, R. STRAUB,
M. WEISHAUPT. 1992. Capillary supply and fibre area in locomotor muscles of
horses and steer- A comparison between histochemistry and electron microscopy,
Acta Anat. 145: 395-399.
KARLSTRÖM, K., B. ESSEN-GUSTAVSSON, S.G.B. PERSSON. 1995.
Capillaries of muscle in red cell hypervolaemic versus normovolaemic standardbred
horses, Equine Vet. J. Suppl. 18: 228-230.
LEIGH, L. 1983. Equinotecnia. Apuntes mimeog. Universidad de
Concepción, Departamento de Medicina Veterinaria, Facultad de Ciencias
Agropecuarias y Forestales, Chillán, Chile.
LINDHOLM, A., K. PIEHL. 1974. Fibre composition, enzyme activity
and concentrations of metabolites and electrolytes in muscles of standardbred
horses, Acta Vet. Scand. 15: 287-309.
LINDHOLM, A., B. ESSEN-GUSTAVSSON, D. McMICKEN, S. PERSSON,
J.R. THORNTON. 1983. Muscle histochemistry of thoroughbred during growth and
training. En: D.H. SNOW, S.G.B. PERSSON and R.J. ROSSE (eds.) Equine Exercise
Physiology. Granta Editions, Cambridge, pp. 211-217.
LOPEZ-RIVERO, J.L., E. AGÜERA, J.G. MONTERDE, M.V. RODRIGUEZ-BARBUDO,
F. MIRO. 1989. Comparative study of muscle fiber type composition in the middle
gluteal muscle of Andalusian, Thoroughbred and Arabian horses, J. Equine
Vet. Sci. 9: 337-340.
LOPEZ-RIVERO, J.L., E. AGÜERA, J.G. MONTERDE, A. DIZ,
J. VIVO. 1990. Fibre size and composition in the middle gluteal muscle of the
Andalusian horse, Equine Vet. J. 22: 286-287.
LOPEZ-RIVERO, J.L., A.L. SERRANO, A.M. GALISTEO. 1992. Variability
of muscle fibre composition and fibre size in the horse gluteus me-dius: An
enzime-histochemical and morphometric study, J. Anat. 181: 1-10.
LOPEZ-RIVERO, J.L., A.M. DIZ, J.G. MONTERDE, E. AGÜERA.
1993. Intramuscular distribution of fibre types in the gluteus medius of the
horse: A histochemical analysis, Anat. Histol. Embryol. 22: 233-240.
LOPEZ, J.L. 1983. Características histoquímicas,
bioquímicas y morfológicas del músculo esquelético
del equino, Agro-Ciencia 9: 113-132.
LOPEZ, J.L., A.M. GALISTEO, A.L. SERRANO, J.L. MORALES. 1993.
Influencia del semental sobre el tipo de composición de fibra muscular
en caballos árabes y andaluces en diferentes estados de desarrollo postnatal
(comunicación preliminar), Agro-Ciencia 9: 43-48.
RIVERO, J.L., M.C. RUZ, A. SERRANO, A.M. DIZ, A.M. GALISTEO.
1993a. Efecto del entrenamiento y desentrenamiento sobre la proporción
de los tipos de fibras musculares en diferentes razas de caballos, Avances
en Ciencias Veterinarias 8: 110-118.
RIVERO, J.L., A.M. DIZ, E. AGÜERA, A.L. SERRANO. 1993b.
Endurance training and muscle fiber types in Andalusian and Arabian horses,
Equine Practice 15: 12-20.
RIVERO, J.L., M.C. RUZ, A. SERRANO, A.M. DIZ. 1995. Effects
of a 3 month endurance training programme on skeletal muscle histochemistry
in Andalusian, Arabian and Anglo-Arabian horses, Equine Vet. 27: 51-59.
RONEUS, M., A. LINDHOLM, A. ASHEIM. 1991. Muscle characteristics
in thoroughbreds of different ages and sexes, Equine Vet. 23: 207-210.
RONEUS, M. 1992. Muscle characteristics in horses in relation
to age, sex and training. Doctoral Thesis. Faculty of Veterinary Medicine, Swedish
University of Agricultural Sciences. Uppsala, Sweden.
RUZ-FERNANDEZ, M.C. 1993. Efectos del entrenamiento, de resistencia
y del desentrenamiento sobre la composición, tamaño y abastecimiento
capilar de los tipos de fibras musculares en diferentes razas de caballos. Tesis
Doctoral, Universidad de Córdoba, España.
SEIFERT, H.S.H. 1971. El caballo de Paso del Perú. Historia,
características y arreos, Noticias Médico Veterinarias
4: 413-440.
SNOW, D.H., P.S. GUY. 1980. Muscle fibre type composition of
a number of limb muscle in differents type of horses, Res. Vet. Sci. 28:
137-144.
SNOW, D.H. 1983. Skeletal muscle adaptations: A review. En:
D.H. SNOW, S.G.B. PERSSON, and R.J. ROSE (eds.) Equine Exercise Physiology.
Granta Editions, Cambridge, pp. 344-353.
VALBERG, S., B. ESSEN-GUSTAVSSON, A. LINDHOLM, S. PERSSON.
1985. Energy metabolism in relation to skeletal muscle fibre properties during
treadmill exercise. Equine Vet. J. 17: 439-444.
VALBERG, S. 1987. Metabolic response to racing and fiber properties
of skeletal muscle in standardbred and thoroughbred horses, Equine Vet. J.
7: 6-12.
VALBERG, S., B. ESSEN-GUSTAVSSON. 1987. Me-ta-bolic response
to racing determined in pools of type I, IIA and IIB fibers. En: GILLESPIE,
J.R., N.E. ROBINSON (eds.) Equine Exercis Physiology 2. ICEEP Publications,
Davis, California, pp. 290-301.
VALBERG, S., B. ESSEN-GUSTAVSSON, A. LINDHOLM, S.G.B. PERSSON.
1989. Blood chemistry and skeletal muscle metabolic responses during and after
different speeds and durations of trotting, Equine Vet. J. 21: 91-95.