ARTICULOS ORIGINALES
SUMMARY
Annual variation in the distribution of bovine trichostrongyle infective
larvae
on pasture grass in Valdivia, Chile
The object of this study was to determine the section of grass
which infective trichostrongyle bovine larvae (L3) prefer and whether larvae
have a daily vertical migration on the grass. Weekly grass samples from a
0.6 ha pasture with calves, from a farm near Valdivia, Chile (39°, 48'S.;
73° 14'W.), were taken from September 1994 (spring) to September 1995
(end of winter). Parasite genera found on the grass samples were Nematodirus
(74.9%), Ostertagia (7.2%), Cooperia (7.0%) and Trichostrongylus (0.4%), without
important seasonal variations. 10.5% of the larvae were not identified
No differences were observed between the weekly samplings of larvae per kg
dry grass (L/kg dg) in the upper (230 L/kg dg) or lower (248 L/kg dg) sections
of grass (p>0.05), during spring and summer. During autumn and winter, the
mean amount of larvae found, was significantly higher in the upper (1.322
L/kg dg) than in the lower (223 L/kg dg) section of the grass (p>0.05). The
highest number of L3 was found in autumn, with great fluctuations and a decreasing
tendency during winter. In 92.8% of the samples, there was no difference in
the amount of larvae on grass collected in the morning (at sunrise) and in
the afternoon, (p<0.05), indicating that there is no vertical daily migration
of the larvae.
Palabras claves: Trichostrongylidae, larvas, ubicación, pasto, potrero, terneros
Key words: Trichostrongylidae, larvae, distribution, grass, calves pasture
INTRODUCCION
El parasitismo gastrointestinal es reconocido como la enfermedad más
común del bovino. Produce dolor abdominal, respuestas anatómicas
y químicas a la infección, cambios en la digestión proteica
y la disponibilidad de aminoácidos absorbibles, cambios en los niveles
de hormonas plasmáticas (gastrina, colecistoquinina) y alteración
del paso de la ingesta. Ello se traduce en anorexia de los animales afectados
con retardo del crecimiento y de la madurez sexual y, en algunos casos, se puede
presentar hasta muerte (Holtenius y col., 1983; Symons,
1985; Rossanigo y col., 1992). Según Entrocasso
(1988), las pérdidas económicas provocadas por los parásitos
gastrointestinales en el ganado bovino de carne argentino ascienden de 20 a
30% de la producción.
En el sur de Chile (X Región), los nemátodos parásitos
más trecuentes del bovino pertenecen a los géneros Cooperia,
Ostertagia y Nematodirus, siendo menos común el género
Trichostrongylus (Oberg y col., 1974; Sievers,
1982; Valenzuela y col., 1992). Estos parásitos,
que afectan preferentemente a los terneros, tienen un ciclo de vida directo
cuyo comportamiento de las larvas infectantes sobre el pasto todavía
es desconocido y es el objetivo de este estudio.
El desarrollo en el medio ambiente se inicia en el momento en que los huevos
de los parásitos caen a la superficie de pastoreo junto con la materia
fecal del animal. Si las condiciones ambientales lo permiten, se desarrollan
larvas denominadas larvas 1 (LI), que eclosionan en la materia fecal y se alimentan
de los elementos allí existentes hasta mudar a larvas 2 (L2); éstas
se siguen alimentando y crecen para culminar su desarrollo con la muda a larvas
3 (L3), que son los estados infectantes. El tiempo que tarda el desarrollo de
huevo hasta L3 varía de una a seis semanas y depende de la temperatura
ambiental y, por ende, de la época del año (Steffan
y Fiel, 1986). Las L3 poseen una cutícula que les impide alimentarse
pero que les confiere gran resistencia frente a las condiciones ambientales,
sin restarles movilidad (Nary y Fiel, 1994). La excepción
a este desarrollo lo presenta el género Nematodirus, en que el
desarrollo a L3 se efectúa dentro del huevo.
Las L3 encuentran en la materia fecal un excelente medio para protegerse de
condiciones climáticas adversas, pero para tener la posibilidad de ser
ingeridas por un hospedador susceptible deben trasladarse al pasto. Dicha traslación
es facilitada, casi exclusivamente, por lluvias fuertes. Las L3 suben a la superficie
de la materia fecal una vez reblandecida la corteza y se ubican en los pequeños
charcos que allí se forman. Las gotas grandes de lluvia torrencial "salpican"
las larvas hacia el pasto hasta una distancia de 60 cm (Gronvold,
1987; Gronvold y Hogh-Schmidt, 1989). Sobre el pasto las
L3 poseen gran movilidad, pero ésta se expresa sólo si existe
suficiente humedad (Armour, 1980; Steffan y Fiel,
1986).
(Nary y Fiel, 1994) señalan que "las larvas infectantes
no se distribuyen homogéneamente a lo largo del pasto. Si bien es cierto
que la mayor concentración se encuentra entre el nivel del suelo y los
10 cm. de altura, esto no es constante, pues las L3 migran activamente en función
de la humedad que tiene la planta. Además, responden en forma inversa
a la intensidad lumínica, de manera que resultaría lógico
encontrar larvas a mayor altura a la salida o la entrada del sol, y los días
nublados y lluviosos. Por el contrario, al progresar el día, cuando la
radiación solar seca el rocío, es probable que las larvas no progresen
en su avance vertical y permanezcan en la broza formada por restos vegetales
y depositada sobre el suelo". Estas suposiciones sobre el comportamiento de
las L3 infectantes de los tricostron-gilidos del bovino se basan en el trabajo
de Williams y Bilkovich (1973) y se citan en muchos textos
de consulta. En este trabajo se ha tratado de revisar dichos conceptos porque
el bovino es una especie de hábitos alimenticios diurnos que arranca
preferentemente la porción superior del pasto y es poco probable que
las L3 eviten ser ingeridas por sus hospedadores, permaneciendo en la parte
inferior del pasto durante el día. La supervivencia de las L3 en el ambiente
está condicionada a sus reservas energéticas, a la temperatura
y a la humedad ambiental (Pandey, 1972). Temperaturas altas
aceleran la movilidad de las larvas y, por ende, éstas consumen sus reservas
energéticas disminuyendo su tiempo de supervivencia (Suárez,
1993). En cambio, temperaturas bajas retardan su metabolismo y, por ello,
viven un tiempo más prolongado (Stetfan y Fiel, 1986).
Por ello es poco probable que las L3, con reservas energéticas limitadas,
tengan una desgasta-dora migración diaria sobre el pasto.
Los objetivos del presente trabajo fueron determinar en qué porción
del pasto se ubican preferentemente las L3 y si éstas tienen migración
diaria vertical ascendente y descendente sobre el mismo.
MATERIAL Y METODOS
El trabajo se realizó en el "Fundo El Bayo", localizado cerca de la
ciudad de Valdivia (39°, 48'S.; 73° 14'W.), X Región, Chile.
Se comenzó en la primavera de 1974 y concluyó al inicio de la
primavera siguiente. Se utilizó un potrero de 0.6 hectáreas sobre
el cual se mantuvieron bovinos menores de un año de edad durante el otoño
e invierno de 1994. A partir de octubre de 1994 y durante todo el ensayo, no
se manejaron bovinos sobre el potrero.
Cada semana se muestreó el pasto del potrero según Kloosterman
(1971), caminando sobre sus diagonales con detenciones cada 10 pasos para
obtener cuatro pequeñas muestras de pasto. Cada pequeña muestra
se dividió en una porción superior y una inferior del pasto, obteniéndose
así, dos muestras acumulativas por cada diagonal del potrero. Las muestras
acumulativas se procesaron mediante la técnica descrita por Sievers
(1973) para obtener la relación larvas por kilogramo de pasto seco
(L/kg ps). Las larvas se diferenciaron según la clave de Bürger
y Stoye (1968). Durante la primavera de 1994 (semanas 1 a 13) y el verano
de 1995 (semanas 14 a 27) se muestreó el pasto dos veces al día,
en la madrugada al salir el sol y en la tarde alrededor de las 15 hrs. Durante
el otoño y el invierno de 1995 (semanas 28 a 52) se muestreó el
pasto sólo en la tarde.
Para determinar si existía diferencia entre las cantidades de larvas
encontradas en las porciones superior e inferior del pasto y entre las cantidades
encontradas en la madrugada y la tarde se utilizó el Test de Kruskall-Wallis
(p<0.05).
La cantidad, en gramos, de pasto seco por metro cuadrado de potrero se estimó
cortando todo el pasto que se encontraba dentro de un anillo de 0.25 m2 arrojado
cuatro veces al azar sobre el potrero. El pasto se secó durante 24 horas
a 60°C en una estufa y luego se pesó mediante una balanza de precisión.
Los datos climáticos se obtuvieron del Instituto de Geociencias de la
Universidad Austral de Chile, situado a 3 km del lugar en estudio. Se calcularon
para cada semana, los promedios de temperatura y la lluvia caída.
RESULTADOS
La relación porcentual de los géneros de parásitos gastrointestinales diagnosticados en las muestras de pasto fue Nematodirus (74.9%), Ostertagia (7.2%), Cooperia (7.0%) y Trichostrongylus (0.4%). Los porcentajes encontrados en las cuatro estaciones del año presentaron variaciones tendiendo a aumentar el género Nematodirus (cuadro 1). No se pudo diferenciar un 10.5% de las larvas debido a que la observación microscópica en cámaras de recuento no permite mover larvas enrolladas o con sus extremos cubiertos por otras partículas.
|
Porcentajes de los géneros de larvas infectantes
encontrados en el pasto de un Percentage of genera of infective larvae found in grass
samples from a pasture with calves |
|||||
|
|
|||||
|
%
|
|||||
| Géneros |
|
||||
| parasitarios |
Primavera
|
Verano
|
Otoño
|
Invierno
|
Promedio
|
|
1994
|
1995
|
||||
|
|
|||||
|
Nematodirus |
77.1
|
71.8
|
71.6
|
79.3
|
74.9
|
| Ostertagia |
6.5
|
6.4 | 11.0 | 4.8 | 7.2 |
| Cooperia | 11.5 | 8.4 | 4.2 | 3.7 | 7.0 |
| Trichostrongylus | 0.0 | 1.4 | 0.1 | 0.0 | 0.4 |
| Indiferenciadas | 4.9 | 12.0 | 13.1 | 12.2 | 10.5 |
|
|
|||||
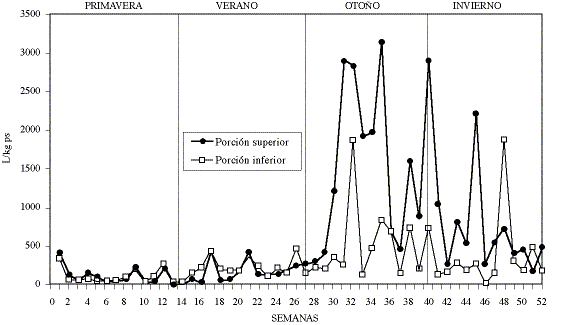
Las cantidades y tendencias de L3 encontradas en la porción superior e inferior del pasto (gráfico 1) son similares (p>0.05) en un 85.2% de los muestreos de primavera y verano, con promedios de 248 y 230 L/kg ps respectivamente. Pero durante otoño e invierno, las L3 se encontraron preferentemente en la porción superior del pasto, existiendo una diferencia significativa (p<0.05) en 82.1% de los muestreos. Los promedios encontrados para otoño e invierno fueron 1.312 y 443 L/kg ps para las porciones superior e inferior del pasto respectivamente. El mayor número de larvas en el pasto se encontró durante el otoño, presentando una tendencia a disminuir durante el invierno. Las cantidades y tendencias de L3 encontradas durante la madrugada y la tarde, a lo largo de la primavera de 1994 y el verano de 1995 (gráfico 2), no difirieron estadísticamente (p>0.05) en un 92.8% de los muestreos.
Gráfico 1. Promedios de larvas por kilogramo de pasto seco (L/kg
ps) encontrados en las porciones superior e inferior de pasto de un potrero
para terneros en Valdivia, Chile, desde septiembre de 1994 a septiembre de 1995.
Average number of infective larvae per kg of dry grass (L/kg dg) found in the
upper and lower section of the grass on a pasture with calves from September
1994 to September 1995, in Valdivia, Chile.
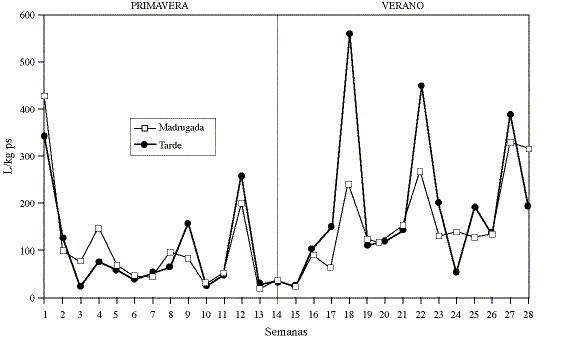
Gráfico 2. Promedios de larvas por kilogramo de pasto seco (L/kg
ps) encontrados en el pasto de un potrero para terneros en Valdivia, Chile,
en la madrugada y en la tarde durante la primavera de 1994 y el verano de 1995.
Average number of infective larvae per kg of dry grass (L/kg dg) on a pasture
with calves at sunrise and in the afternoon, during spring 1994 and summer 1995
in Valdivia, Chile.
La cantidad de materia seca de pasto alcanzó en promedio 680 g/m2 durante
la primavera de 1994 y disminuyó paulatinamente a 150 g/m2 durante el
invierno de 1995.
Los datos climáticos de cada semana se presentan como promedio de temperatura
y suma de lluvia caída (gráfico 3). Durante
la primavera de 1994 la temperatura promedio semanal fue aumentando hasta alcanzar
los 18°C durante el verano de 1995; durante el otoño disminuyó;
presentando un mínimo de 2.9°C en la tercera semana del invierno
de 1995, aumentando posteriormente en forma gradual. Durante la primavera y
el inicio del verano hubo lloviznas de cierta intensidad, pero las lluvias fuertes
se concentraron en los meses de otoño e invierno, alcanzando un máximo
de 245 mm en la última semana del otoño.
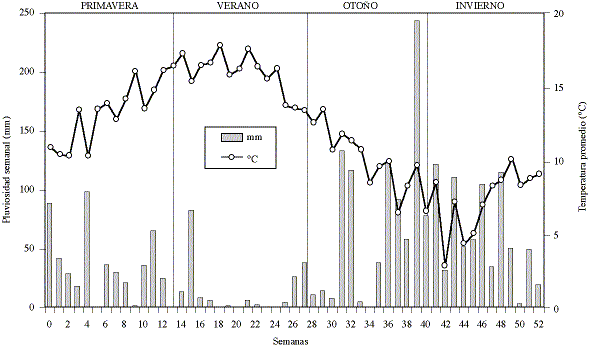
Gráfico 3. Temperatura promedio y lluvia semanal de septiembre
de 1994 a septiembre de 1995 en Valdivia, Chile.
Mean weekly temperature and rainfall from September 1994 to September 1995 in
Valdivia, Chile.
DISCUSION
El predominio del género Nematodirus frente a Ostertagia
y Cooperia (cuadro 1) no concuerda con lo observado
para la región (Oberg y col., 1974; Valenzuela
y col., 1992), y sólo se puede explicar dicha diferencia en forma
especulativa por el régimen de desparasitaciones implantado en los últimos
años por el agricultor, en que todos los bovinos menores de un año
se trataban una vez al mes con el fármaco Ivermectina. Como el producto
no es 100% eficiente contra Nematodirus (Robles, 1983),
es probable que la baja eliminación de huevos de este parásito
haya provocado la contaminación predominante del potrero.
El número total de L3 en el pasto fue regularmente bajo durante la primavera
1994 y el verano 1995. En cambio en otoño de 1995 aumentó cerca
de 10 veces el número de larvas (gráfico l).
Las precipitaciones que hubo durante la primavera y el inicio del verano, si
bien fueron abundantes, tuvieron características de llovizna y, al parecer,
no fueron suficientes para trasladar las L3 al pasto. El enorme aumento del
número de larvas sobre el pasto en otoño se debió, con
toda seguridad, a su traslación masiva desde la materia fecal al pasto,
favorecida por las lluvias torrenciales que se presentaron a partir de la semana
28 (gráfico 3). Ello corrobora, para la realidad
nacional, lo descrito por Gronvold (1987) y Gronvold
y Hogh-Schmidt (1989) en relación a la participación de las
lluvias en la traslación de las L3 de la materia fecal al pasto. Sin
embargo, es necesario agregar que, después de lluvias torrenciales, se
presentaron drásticas disminuciones del número de L3, lo que se
puede deber a un efecto de lavado del pasto con arrastre de larvas hacia el
suelo. Posteriormente, por la humedad presente en el pasto, fue posible la migración
activa ascendente de las L3 y una pronta recuperación de su número
en la porción superior del pasto.
La última contaminación del potrero con huevos de parásitos
se produjo en la primera semana de octubre de 1994 (primavera). De los huevos
se desarrollaron L3 que permanecieron en gran medida cautivas en la materia
fecal hasta abril de 1995 (otoño), momento en que las lluvias torrenciales,
con gotas grandes, facilitaron su traslación masiva al pasto. Gran parte
de dichas L3 sobrevivieron sobre el pasto hasta la primavera siguiente en una
cantidad suficiente como para infectar una nueva generación de terneros.
Las observaciones coinciden con Steffan y Fiel (1986) y Entrocasso
(1988) que describen que en primavera y verano las deposiciones fecales
se convierten en reservorios de larvas hasta el otoño siguiente, permitiendo
su sobrevida por más de 18 meses, facilitando así el pasaje de
la infección de un ciclo de producción de una temporada a otra.
Durante primavera y verano no existió diferencia entre la cantidad de
L3 encontradas en la porción superior e inferior del pasto, pero durante
otoño e invierno fue mayor la cantidad de L3 en la porción superior
(gráfico l). Ello difiere de lo señalado
por Nari y Fiel (1994), que describen que las L3 se ubican
mayoritariamente en la parte inferior del pasto. Por ello se plantea la hipótesis
de que se trata de una adaptación evolutiva de los parásitos del
bovino, que debe favorecer su posibilidad de ser ingeridos, ya que esta especie
arranca preferentemente la porción superior del pasto y sólo come
a ras del suelo cuando la disponibilidad de forraje es escasa.
No hubo diferencia entre las cantidades de L3 encontradas sobre el pasto en
la madrugada y en la tarde, durante la primavera de 1994 y el verano de 1995
(gráfico 2), lo que indica indirectamente que las
L3 no tienen migración diaria ascendente y descendente sobre el pasto,
estando en desacuerdo con lo citado por Nari y Fiel (1994).
Durante otoño e invienio de 1995 se obtuvieron las muestras sólo
en la tarde, supuestamente en el momento en que debieran haber menos larvas
en la porción superior del pasto y, sin embargo, su cantidad fue regularmente
mayor (gráfico 1).
Se debe tener en cuenta que las L3 no se alimentan y que basan su supervivencia
en las reservas energéticas adquiridas como L1 y L2. El realizar migraciones
diarias desde la base hasta las partes altas del pasto, significaría
para las L3 un gasto energético que reduciría sus posibilidades
de supervivencia. Por este motivo es más probable que las larvas migren
sólo en dirección ascendente, tratando de permanecer siempre en
la porción superior del pasto. Las enormes fluctuaciones en las cantidades
de L3 sobre el pasto (gráfico 1) indican que las
disminuciones y aumentos podrían deberse al arrastre de las L3 hacia
el suelo por efecto de las lluvias y la migración activa ascendente de
las L3 que tratan de ubicarse donde puedan ser ingeridas por un bovino.
Los datos climáticos obtenidos durante el ensayo (gráfico
3) concuerdan, en gran medida, con lo descrito por Huber (1970)
para Valdivia, en que la precipitación promedio anual fluctúa
entre los 2.200 a 2.700 mm, con un período "lluvioso" entre abril y septiembre
y un período relativamente "seco" entre octubre y marzo. Esta condición
macroclimática permitió que la variación anual de la cantidad
de pasto del potrero estudiado también correspondiera a lo esperado.
Se concluye que durante primavera y verano las L3 se encuentran en cantidades
bajas, tanto en la porción superior como inferior del pasto. En cambio,
durante el otoño aumenta en 10 veces su cantidad, ubicándose preferentemente
en la porción superior del pasto. Durante el invierno las L3 también
se ubican en la porción superior pero su número tiende a disminuir.
No hubo diferencia entre la cantidad de larvas encontradas sobre el pasto en
la madrugada y en la tarde de primavera y verano, lo que indicaría que
no hay una migración diaria ascendente y descendente sobre él.
RESUMEN
El trabajo se realizó con muestreos semanales de pasto de un
potrero de 0.6 has. destinado a terneros en un predio cercano a la ciudad
de Valdivia (39°, 48' S.; 73° 14' W.), Chile, desde septiembre
de 1994 (primavera) a septiembre de 1995 (fines de invierno). El objetivo
fue determinar en qué porción del pasto se encuentran preferentemente
las larvas infectantes (L3) de los tricostrongilidos del bovino y si éstas
tienen migración diaria ascendente y descendente sobre el pasto.
Los géneros de parásitos diagnosticados en las muestras de pasto
fueron Nematodirus (74.9%), Ostertagia (7.2%), Cooperia
(7.0%) y Trichostrongylus (0.4%). No se diferenció un 10.5% de
las larvas.
Durante primavera y verano no hubo diferencia (p<0.05) entre las
cantidades de L3 encontradas en las porciones superior e inferior del pasto
en un 82.1% de los muestreos con promedios de 230 y 248 L/kg ps, respectivamente.
Durante otoño e invierno hubo diferencia (p>0.05) entre ambas porciones
en un 82.1% de los muestreos, con promedios de 1.312 y 443 L/kg ps, en
las porciones superior e inferior del pasto, respectivamente. El mayor
número de L3 se encontró durante el otoño con grandes
fluctuaciones y con una tendencia a disminuir a lo largo del invierno.
Entre las cantidades de L3 encontradas en el pasto en la madrugada y la
tarde no hubo diferencia (p<0.05) en 92.8% de los muestreos, lo que
indica que no hubo migración ascendente y descendente diaria sobre
el pasto.
AGRADECIMIENTOS
Al señor Enrique Sinning, propietario del predio, por su desinteresada colaboración para realizar el trabajo.
DEDICATORIA
Deseamos dedicar el presente trabajo, con cariño y reconocimiento, al primer profesor de Parasitología de la Facultad de Ciencias Veterinarias de la Universidad Austral de Chile, Prof. Dr. José S. Tolosa Palacios.
Aceptado: 16.10.97.
BIBLIOGRAFIA
ARMOUR, J. 1980. The epidemiology and control of helminth disease in farm animals, Vet. Parasitol. 6: 7-46.
BÜRGER, H.J., M. STOYE. 1968. Parasitologische Diagnostik. Eizählung und Larvendifferenzierung, München. Therapogen Praxisdienst (Inf. Técnico 24).
ENTROCASSO, C M. 1988. Epidemiology and control of bovine ostertagiasis in South America, Vet. Parasitol. 27: 59-65.
GRONVOLD, J. 1987. A field experiment on rain splash dispersal of infective larvae of Ostertagia ostertagi (Trichostrongylidae) from cow pats to sorrounding grass, Acta Vet. Scand. 28: 459-461.
GRONVOLD, J., K. HOGH-SCHMIDT 1989. Factors influencing rain splash dispersal of infective larvae of Ostertagia ostertagi (Trichostrongylidae) from cow pats to the sorroundings, Vet. Parasitol. 31: 57-70.
HOLTENIUS, P., A. UGGLA, G. OLSSON. 1983. Epidemiological studies on Ostertagia ostertagi infections in cattle during their first, second and third summer on pasture, Nord. Vet. Med. 35: 233-238.
HUBER, A. 1970. Diez años de observaciones climatológicas en la estación Teja-Valdivia (Chile) 1960-1969. Universidad Austral de Chile, Facultad de Ciencias Naturales y Matemáticas Instituto de Geología y Geografía.
KLOOSTERMAN, A. 1971. Observations on the epidemiology of trichostrongylosis of calves. Veenan and Zoonen, N. V. Wageningen.
NARI, A., C. FIEL, 1994. Enfermedades parasitarias de importancia económica en bovinos. Bases epidemiológicas para su prevención y control. Editorial Agropecuaria Hemisferio Sur, Montevideo, p. 76.
OBERG, C., L. DIAZ, G. VALENZUELA. 1974. Parásitos identificados en bovinos, ovinos, suinos y equinos en el laboratorio de Enfermedades Parasitarias de la Escuela de Medicina Veterinaria de la Universidad Austral de Chile, 1963-1973. Bol. Chile Parasit. 29: 99-102.
PANDEY, V.S. 1972. Effect of temperature on survival of the free-living stages of Ostertagia ostertagi, J. Parasitol. 58: 1042-1046.
ROBLES, S.A. 1983. Efecto del fármaco ivermectina (IVOMEC) sobre la eliminación de huevos de parásitos gastrointestinales en las fecas de terneros en sus primeros meses de pastoreo. Tesis, M.V., Universidad Austral de Chile, Facultad de Ciencias Veterinarias, Valdivia, Chile.
ROSSANIGO, C.E., J.D. AVILA, R.L. SAGER 1992. Parasitismo gastrointestinal subclínico en terneros de destete: su efecto sobre el consumo, la digestibilidad y la ganancia de peso, Rev. Med. Vet. 73: 88-92.
SIEVERS, G. 1973. Methode zur Gewinnung von III. Strongylidenlarven aus dem Weidegras. Tesis de doctorado. Tierärztliche Hochschule, Hannover, República Federal de AIemania.
SIEVERS, G. 1982. Epizootiología de las trichos-trongilidosis de los terneros en Chile. En: VIII Jornadas Médico-Veterinarias, 26, 27 y 28 de agosto de 1982, Valdivia, Chile, pp 93-112.
STEFFAN, P.E., C.A. FIEL. 1986. Bioecología de los nemátodos gastrointestinales de los bovinos, Rev. Asoc. Arg. Prod. Anim. 6: 139-140.
SUAREZ, V.H. 1993. Epidemiología de los nemátodos de la región subhúmeda y semiárida pampeana. Enfermedades parasitarias de importancia económica en bovinos. Editorial Hemisferio Sur, Buenos Aires.
SYMONS, L.E.A. 1985. Anorexia. Occurrence, pathophysiology, and possible causes in parasitic infections, Adv. Parasitol. 24: 103-133.
VALENZUELA, G., G SIEVERS, E.I. QUlNTANA 1992. Parásitos identificados en bovinos, ovinos y equinos en el Laboratorio de Parasitología de la Facultad de Ciencias Veterinarias de la Universidad Austral de Chile. 5ª Jorn. An. Parasitol., Olmué, Chile, pp 43-45.
WILLIAMS, J.C., F.R. BILKOVICH. 1973. Distribution of Ostertagia
ostertagi infective larvae on pasture herbage, Am. .J. Vet. Res. 34:
1337-1334.