P. ORELLANA1, M.V. M. Sc.; N. MENDOZA2, Q.F. M.S.; M. SCORI, M.V.
1Nutrición Animal, Depto. de Cs. Pecuarias,
Facultad de Medicina Veterinaria, Universidad de Concepción,
Casilla 537, Chillán, Chile. 2Bromatología,
Depto. de Nutrición y Dietética, Facultad de Farmacia,
Universidad de Concepción, Chile.
SUMMARY
Relationship between urinary excretion of purine and creatinine derivatives and food intake in dairy cows
Molar concentrations of purine (PD) and creatinine (CT) derivatives were measured in urine samples of 63 cows submitted to 4 levels of feeding (6.04; 10.09; 11.63 y 18.31 kg DM/d and 2.98; 5.9; 7.37 y 12.5 kg DOMR/d). Based on PD/CT ratio and dry matter intake (DMI) and degradable organic matter in the rumen, intake (DOMRI), regression equations were derived to estimate of DOMRI based on the PD/CT ratio.
Moreover, the daily excretion of PD was calculated and from that, the absortion of PD and efficiency of microbial nitrogen supply (EMNS) by food intake unit were obtained.
It was concluded that the molar PD/CT ratio in one urine sample, allows to discriminate among levels of food intake differing more than 4.05 kg of DMI, or more than 2.92 kg of DOMRI.
The quadratic regression were PD/CT = 0.25 + 0.072 DMI + 0.002 DMI2 (r2 = 0.720) and PD/CT = 0.25 + 0.147 DOMRI + 0.0015 DOMRI2 (r2 = 0.710 ). However, when they were expressed as the total of PD excreted daily (PDe), it was possible to differentiate among the 4 levels of feeding. Values of PDe by groups were I = 84.77 ± 12.19; II = 140.66 ± 7.21; III = 168.67 ± 6.45 and IV = 297.03 ± 9.72 (mmol/d) (p< 0.05).
The ± 0.928; II = 8.096 ± 0.549; III = 8.898 ± 0.491 and IV=11.573 ± 0.74 g/kg where the group IV was the only one different from the others (p< 0.05). Expressed as EMNS/kg of DOMRI was statistically equal for the 4 groups with values of: I = 13.224 ± 1.537; II = 13.846 ± 0.909; III = 14.041 ± 0.813 and IV = 16.953 ± 1.226 g/kg respectively. It is concluded that PD/CT of a urine sample in a cow has a high correlation with food intake and that PDe estimated from this ratio allows to identify differences in average food intake of 1.5 kg of DMI and of 1.47 kg of DOMRI
Palabras claves: derivados de purinas, creatinina, nitrógeno microbiano, vacas de lechería
Key words : purine derivatives, creatinine, microbial nitrogen, dairy cows.
INTRODUCCION
Los derivados de purinas (DP) excretados en la orina de rumiantes incluyen a la alantoína (A), ácido úrico (AU) xantina (X) e hipoxantina (HX). Estos metabolitos se originan de la degradación de las purinas, tanto de origen endógeno como exógeno. En los rumiantes las purinas exógenas se originan casi exclusivamente de los ácidos nucleicos de los microorganismos ruminales que son metabolizados por el animal, excretándose principalmente como alantoína (Fujihara, 1987).
Numerosos trabajos han demostrado que existe una alta correlación entre la infusión abomasal de ácidos nucleicos con la excreción de los DP urinarios. (Giesecke y col., 1984; Linberg y col., 1989; Chen y col., 1990; Verbic y col., 1990; Bacells y col., 1991). También se ha demostrado una alta correlación entre la excreción de los DP con la infusión abomasal de microorganismos exógenos (Verbic y col, 1990) y con el consumo de alimento en vacas (Gonda y col., 1994), en cabras (Lindberg, 1985) y en ovejas (Chen y col., 1992b).
La determinación de los DP urinarios es un indicador útil en estudios de nutrición de rumiantes, ya que a partir de ellos es posible calcular la cantidad de ácidos nucleicos microbianos metabolizados por el animal obtenidos de los microorganismos ruminales (MO). Por medio de la excreción de los DP y de la tasa en que se encuentra el nitrógeno de las purinas con respecto al nitrógeno proteico microbiano, es posible estimar la síntesis de nitrógeno proteico microbiano que se realiza en el rumen (Chen y Gomes, 1992). Al relacionar la síntesis de nitrógeno microbiano con la cantidad de alimento consumido, se obtiene el rendimiento (K) de nitrógeno proteico microbiano, el que normalmente se expresa como: K de nitrógeno microbiano/kg materia seca (NM/MS) o nitrógeno microbiano/kg de materia orgánica (NM/MO) o bien como nitrógeno micro-biano/kg de materia orgánica degradable en el rumen (NM/MODR) (Chen y col., 1992).
La técnica para obtener la totalidad de los DP excretados diariamente requiere la recolección diaria total de orina, pero para poder hacerla aplicable en animales en producción es necesario simplificarla hasta el punto que sólo baste obtener muestras puntuales de orina. Esta simplificación es posible, ya que la producción de los DP es constante durante el día (Chen y col., 1992a) y la dilución de la orina es posible corregirla mediante la concentración de la muestra urinaria con un marcador interno o externo. El marcador (M) debe ser un metabolito que se excrete de manera constante por la orina, que sea independiente tanto del consumo de alimento como de la producción y de la digestión de los MR. Así, es posible expresar la excreción de los DP como la tasa DP/M y a partir de ésta es posible calcular el total de los DP excretados diariamente, Faichney y col. (1995).
La creatinina (CT) es un metabolito que cumple con los requisitos para ser considerada como un marcador interno, ya que se produce a una tasa constante a partir de la fosfocreatina y es distribuida a través del agua corporal (Finco, 1980) y según Brody (1945) es excretada en proporción al peso vivo (N-Creatinina (mg/d) = 12.7 PV0.896) dentro de un amplio rango de pesos (0.02 a 800 kg de PV), es independiente de la dieta (Lindberg y Jacobson, 1990) y se excreta a una tasa constante con un baja variabilidad durante el día (Antoniewicz, 1981; Orskov, MacLeod, 1982a; Lindberg, 1989).
Antoniewicz y col. (1981) encontraron, en ovejas, que la excreción diaria de alantoína (A) y creatinina (CT) fueron lo suficientemente constantes como para predecir la excreción de A a partir de la tasa A/CT de la orina, con una alta correlación con los valores medidos en la orina total (r= 0.86, CV= 13.3%).
En el presente trabajo se puso a prueba la hipótesis de que a partir de la tasa DP/CT de una muestra de orina de vacas es posible discriminar el nivel de alimentación a la que están sometidas.
MATERIAL Y METODOS
Se utilizaron 63 vacas Holstein, provenientes de tres lecherías de la Octava Región de Chile (comuna de Chillán), conformando los siguientes 4 grupos de acuerdo a su nivel de alimentación.
Grupos experimentales y características de las dietas ofrecidas.
|
|
||||
|
|
|
|
MO (%) |
|
|
|
|
MO (%) |
||
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
El número de vacas por cada grupo y su condición de lactancia fue el siguiente:* MSI = materia seca ingerida (kilos/animal/día); MO = materia orgánica; MODR = materia orgánica degradable en el rumen (kilos/animal/día).
** PT = paja de trigo; TB = trébol blanco/ballica; E.M. = ensilaje de maíz; R = remolacha hojas y coronas; M = melazam®; H = heno de alfalfa; Cr = coseta remolacha; C = concentrado.
Número de vacas por grupos y sus
características productivas.
|
|
|||
|
|
vacas |
lactancia |
producción leche (L/día) |
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
Análisis de alimentos. La materia seca (MS) fue determinada por deshidratación de la muestras a 60°C durante 48 hrs y la materia orgánica (MO) se determinó descontando el peso de las cenizas obtenidas por incineración de las muestras a 600 C durante 24 hrs. La degradabilidad de la MO en el rumen (MODR) se realizó in sacco en tres ovejas fistuladas ruminalmente, de acuerdo al método de Orskov, Hovell y Mould (1980). Los forrajes fueron incubados por 48 hrs y los concentrados durante 24 hrs.* 6 a 8 meses de gestación.
Alimentación de los animales. Los animales se mantuvieron confinados en grupos de alimentación de acuerdo a su nivel de producción. El alimento fue ofrecido en cuatro raciones diarias (7, 11, 15 y 19 hrs) durante 5 días (4 días previos y durante el día de la obtención de la muestra) y se determinó diariamente el consumo por grupo, restando al alimento ofrecido el remanente de los comederos (base materia seca). Se asignó como consumo individual el consumo promedio del grupo.
Pesaje de los animales y obtención de muestras de orina. Los animales fueron pesados individualmente mediante una balanza electrónica portátil Ruddweigh KM2-E microcomputarizada, la cual también fue utilizada para pesar el alimento.
Las muestras de orina fueron obtenidas entre las 15 y 16 horas mediante una sonda vesical, extrayéndose aproximadamente 10 ml de muestra, la que fue acidificada con ácido sulfúrico para llevarla a pH < a 3 para evitar la proliferación bacteriana y posteriormente fueron congeladas a -20°C durante un máximo de 30 días.
Análisis de los derivados de purinas y creati-nina. Los análisis de creatinina, ácido úrico y alantoína fueron realizados mediante cromatografía líquida de alta resolución (HPLC) de acuerdo a la técnica descrita por Resines, Arín y Diez (1992), utilizando un Cromatógrafo Hitachi L-500 con bomba programable 655A-11, un detector UV 655A acoplado a un integrador D-2000 y una columna LiChroSpher 100-RP 18 5 µm (244 x 4 mm).
Identificación y cuantificación de los derivados de purinas y creatinina. Se prepararon soluciones patrones con cada una de las sustancias a cuantificar ( Sigma p.a.) con concentraciones entre los 5 a 50 mg/L., las cuales fueron utilizadas para identificar las sustancias mediante los tiempos de retención y para cuantificarlas mediante la regresión entre las áreas de los picos de las muestras con las concentraciones de las soluciones patrón.
Las muestras de orina fueron diluidas 1/100 con solución stock y filtradas (0.45 um) antes de ser inyectadas en el cromatógrafo.
De las muestras de orina de cada animal se obtuvo la concentración molar de los derivados de purinas (DP) y de creatinina (CT), para obtener la tasa DP/CT.
La excreción diaria de los derivados de purinas (DPe) fue calculada mediante la siguiente fórmula, citada por Faichney y col. (1995):
Fórmula 1:
|
DP x (PV x Kct)/113.12
|
|
| DPe = |
|
|
CT
|
DPe = Excreción diaria de los derivados de purinas (mmol/d).
DP = Concentración de los derivados de purinas en la muestra de orina (mmo/L).
PV = Peso vivo (kg).
CT = Concentración de creatinina en la muestra de orina (mmol/L).
KCT = Coeficiente de excreción diario de creatinina (mg/d.K) = 113 PV-0.25 obtenida de los datos citados por Orskov y MacLeod (1982) y Chen y col. (1992a).
113.12 = Peso molecular de la creatinina.
Fórmula 2:
|
DPe - (0.3.85 x PV0.75)
|
|
| PA = |
|
|
|
0.85
|
PA = Purinas absorbidas por día
(mmoles/d).
DPe = Excreción diaria de derivados de purinas (mmoles).
0.85 = Factor de recuperación como DP de las purinas absorbidas.
(0.385 x PV0.75) = Contribución endógena de DP mmoles/por kilo de peso metabólico.
Fórmula 3:
|
PA X 70
|
|
| NM = |
|
|
|
0.83 x 0.116 x 1000
|
NM = Nitrógeno microbiano (g/d).
PA = Purinas absorbidas (mmol/d).
70 = Contenido de nitrógeno en las purinas (mg N/mmol).
0.83 = Factor de digestibilidad de las purinas.
0.116 = Tasa de N de purinas: N total en microorganismos ruminales (11.6: 100).
1000 = Factor de corrección de mg a g.
Análisis estadístico. De las muestras de orina de cada animal se obtuvo la concentración molar de los derivados de purinas (DP) y de creatinina (CT), para obtener la tasa DP/CT con la cual se calcularon las ecuaciones de regresión con el consumo de MS y MODR.
La tasa DP/CT, los DPe el NM y el rendimiento de NM. fueron analizados mediante ANDEVA y el test de Scheffé con el programa SYSTAT 5.04. Se consideró una probabilidad del 95% (p< 0.05) para ser considerado estadísticamente diferente.
RESULTADOS
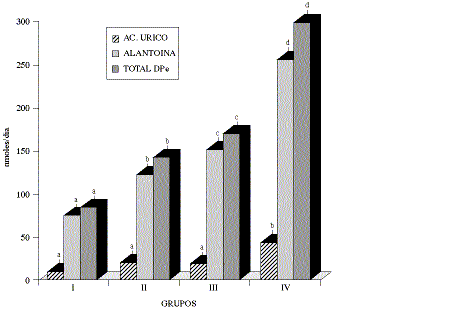
La tasa DP/CT fue diferente sólo entre los grupos que diferían en su alimentación en 4.0 kg MS. y de 2.9 kg de MODR (cuadro 3), mientras que al expresar estos valores como DPe, éstos fueron estadísticamente diferentes entre todos los grupos (gráfico 1).
Las ecuaciones de regresión cuadrática fueron las que mejor relacionaron los valores de los DPe urinarios con el consumo de materia seca (MSI) o MODR, y entre ellas, la que tuvo un mayor coeficiente de determinación fue la relación de los DPe con la MSI (r2=0.800) (cuadro 4), lo que indica que un 80% de la varianza de la excreción de los DPe se debe a su asociación con la variación de alimentación.
*letras diferentes en las mismas columnas entre grupos, indican diferencias estadísticas (p<0.05)
Gráfico 1. Excreción urinaria diaria estimada de
los derivados de purinas (DPe) en vacas, por grupo de tratamiento (![]() mmol/d ± E.E.)
mmol/d ± E.E.)
Estimated urinary daily excretion of purine derivatives (DPe) in cows,
per treatment group ![]() mmol/d ±
S.E.).
mmol/d ±
S.E.).
Las ecuaciones de predicción para estimar el consumo de materia orgánica degradable en el rumen (MODR), a partir de la tasa urinaria de los derivados de purinas/creatinina (DP/CT) y del total de los derivados de purinas excretados diariamente (DPe), se muestran en el cuadro 5 y en ellas se observa que la mejor estimación corresponde a la ecuación que utiliza a los DPe como variable independiente, la cual explica que un 74.5% de la variación del consumo de MODR se debe a su asociación con la excreción de los DPe.
El rendimiento de nitrógeno microbiano (NM) producido por kg de MS sólo fue diferente en el grupo con mayor consumo de materia seca (MSI), el que también corresponde a la dieta con mayor degradabilidad de la materia orgánica (76.05%) debido al mayor contenido de concentrado, pero al expresarlos en base al consumo de materia orgánica degradable en el rumen (MODR) el rendimiento fue igual en todos los grupos con valores entre 13.224 a 16.953 g de NM/kg MODR. (cuadro 6).
Tasa de los derivados de purinas/creatinina
en orina de vacas, por grupo de tratamiento (![]() ± E.E.).
± E.E.).
Ratio of purine/creatinine derivatives
in cow urine per treatment group (![]() ± S.E.).
± S.E.).
|
|
||
|
|
|
|
|
|
||
|
|
|
|
|
|
|
1.212 ± 0.063 b
|
|
|
|
|
|
|
|
|
|
|
||
* Letras distintas en las columnas indican diferencia estadística (p<0.05 test de Scheffé).
= promedio. E.E. = error estándar.
Ecuaciones de regresiones de la excreción diaria de los derivados
de purinas estimados (DPe) y la tasa
derivados de purinas/creatinina (DP/CT) con el consumo de alimento (MSI y
MODR).
Regression equation of estimated daily purine derivatives excretion (DPe) and the ratio purine/creatinine derivatives ratio (DP/CT) with food intake (MSI and MODR).
|
|
|||||||||||
|
Ecuación de regresión |
|||||||||||
|
|
|||||||||||
| DP/CT* |
|
|
|
|
|
|
|
|
|
r2 | = 0.720 |
|
|
|||||||||||
| DP/CT |
|
|
|
|
|
|
|
|
|
r2 | =0.710 |
|
|
|||||||||||
| DPe |
|
|
|
|
|
|
|
|
|
r2 | = 0.800 |
|
|
|||||||||||
| DPe |
|
|
|
|
|
|
|
|
|
r2 | =0.798 |
|
|
|||||||||||
|
* DP/CT = tasa de la concentración de los derivados de
purinas y creatinina urinaria (mmoles). |
|||||||||||
Ecuaciones de regresión predictivas del consumo de materia orgánica degradable en el rumen (MODR).
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
| Valores entre paréntesis = error de la estimación para un 95% de seguridad | |||||||
Rendimiento de nitrógeno microbiano estimado, por unidad de materia
seca y de materia orgánica
degradable en el rumen (g/kg) (![]() E.E.).
E.E.).
|
|
||
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
| * | NM/MSI = nitrógeno microbiano producido (g) por kilo de materia seca ingerida. | |
| ** | NM/MODR = nitrógeno microbiano
producido (g) por kilo de materia orgánica digestible en el rumen. Letras supercriptas diferentes en las columnas indican significancia estadística (p<0.05 test de Scheffé). |
|
DISCUSION
Los promedios de la tasa DP/CT de una muestra de orina permitieron diferenciar a qué grupos de alimentación pertenecen las vacas, cuando las diferencias de consumo por grupo fueron mayores de 4.0 kg MS. y de 2.9 kg de MODR. La ecuación de regresión cuadrática presentó un alto coeficiente de regresión (r2= 0.720 y 0.710) con el consumo de MSI y MODR, respectivamente. Estos valores son levemente inferiores a la regresión lineal calculada por Vercoe (1976), quien obtuvo un r2 = 0.81, entre el consumo de materia seca digestible y la excreción de alantoína, al utilizar novillos con alimentación controlada individualmente, con consumos entre 1 a 5 kg de materia orgánica (MO) (58 a 60% de digesti-bilidad de la MS). Pero los presentes resultados fueron superiores a los reportados por Lindberg (1985) en cabras, quien obtuvo un r2= 0.689 entre la excreción de alantoína y la materia orgánica digestible ingerida.
Esto indica que la tasa promedio de DP/CT del grupo, obtenidas de muestras puntuales de orina, puede ser considerado como un buen indicador del consumo promedio grupal.
Al corregir los valores de la tasa DP/CT por el peso vivo (PV) y por el coeficiente de excreción diario de creatinina, según la fórmula de Faichney y col., (1995), y utilizando el coeficiente de excreción de creatinina (113 PV-0.25) calculado de los datos entregados por Orskov y MacLeod (1982a) y Chen y col. (1992a), se calculó el total de los derivados de purinas excretados en el día (DPe), resultando este parámetro un mejor indicador del consumo grupal, ya con él se pudo diferenciar entre grupos de alimentación, con diferencias de consumo de 1.54 kg de MS y 1.47 kg de MODR.
Las ecuaciones que relacionan el consumo de MSI o de MODR con los DPe tuvieron un r2= 0.8 y 0.798, respectivamente, valores que son superiores a la ecuación de predicción encontrada por Lindberg (1985) en 14 cabras con alimentación controlada individualmente, cuya ecuación predictiva de la excreción de alantoína tuvo un r2 = 0.739 con los valores observados.
El rendimiento de producción de NM/kg MSI fue estadísticamente mayor sólo en el grupo IV, lo que se debería a su mayor proporción de concentrado, pero al expresarlo por kg de MODR no hubo diferencias entre los grupos, encontrándose valores entre 13.224 ± 1.536 y 16.953 ± 1.226 g NM/kg MODR, los cuales son inferiores a los 30 g NM/kg MODR asumidos por el ARC (1984), pero similares a los encontrados por Susmel y col., (1994) en vacas Simmental con alimentación individual, cuyos valores fueron de 12.25 g N/kg MODR para una dieta de 6.15 kg MS en base a paja de trigo (5.44 kg), complementada con afrecho de soya (0.71 kg), dieta que fue similar a la del grupo I, mientras que el mejor rendimiento (18.5 g N/kg MODR) lo obtuvieron con 7.68 kg de heno de festuca más 2.65 kg de harina de maíz, dieta equivalente en cantidad a la del grupo III, en la que se obtuvo un rendimiento de 14.04 g NM/kg MODR. Estos valores también son inferiores a los encontrados por Chen y col. (1992a) en novillos (27.1 a 29.9 g NM/kg MOD) y también a los reportados por Chen y col. (1992b) en ovejas (12.0 a 28.3 g NM/kg MODR). Estas diferencias pueden deberse a que la eficiencia de utilización de los alimentos está en relación al tamaño de los animales, como fue demostrado por Chen y col. (1992b) en ovejas, las que al estar consumiendo la misma cantidad y calidad de dieta, la eficiencia de producción de MM varió de 8 a 36 g NM/kg MODR, ambos relacionados in-versamente con el peso vivo de los animales. Ellos explican este fenómeno a que la tasa CMS/PV (consumo de materia seca/peso vivo) define la tasa de pasaje y por lo tanto el eflujo de MM desde el rumen al intestino. El similar peso de las vacas utilizadas por Susmel y col. (1994) hace más comparables los resultados de dicho estudio con los obtenidos en el presente trabajo.
Es notorio el relativamente alto valor del rendimiento de NM/MODR en los animales con la dieta I en base a paja de trigo, la que a pesar de tener la menor digestibilidad de la MO (54.61%) no fue estadísticamente diferente con el resto de las dietas, las que tenían digestibilidades entre 69.05 y 76.05%.
Esto puede ser explicado por la alta tasa ATP/biomasa microbiana encontrada en dietas de este tipo por Susmel y col. (1991 y 1994) y también por Wolstrup y Jensen (1976), Forberg y Lam (1977) y Erfle y col. (1979). Esta alta concentración de ATP puede deberse a que en dietas con carbohidratos de lenta degradación las bacterias harían una mejor utilización de la energía que aquellas que contienen sustratos más rápidamente fermentables o con una mayor tasa de pasaje de alimentos picados o molidos como lo fue en las dietas III y IV.
Por otra parte, la variabilidad en la excreción de DPe se debería a que la tasa microbiana de N-ARN/N total, asumida como 0.116, parece no ser siempre constante y, según algunos autores, ésta puede variar durante el crecimiento bacteriano (Arambel y col., 1982; Bates y col., 1985 y Susmel y col., 1993), lo que puede causar una variabilidad en la excreción de los DP no relacionados directamente con la biomasa microbiana.
Las ecuaciones de regresión entre la excreción de DP/CT y de DPe con respecto al consumo de alimentos (MSI y MODR) tuvieron un alto coeficiente de regresión, superior al obtenido por Vercoe (1976) en bovinos con recolección total de orina y con alimentación controlada individualmente, quien obtuvo una ecuación de regresión lineal con un r2 = 0.651 entre la excreción de alantoína y el consumo de materia seca digestible, ello probablemente debido al menor número de animales utilizados en dicho estudio (6 animales con dos dietas y 3 niveles de alimentación cada una).
De los resultados obtenidos se concluye que la excreción diaria de derivados de purinas (DPe) obtenidos de la tasa DP/CT determinados en muestras de orina de vacas, presentan un alto coeficiente de regresión con el consumo de alimento, lo que permite discriminar diferencias de consumo promedio de 1.54 de MSI y de 1.47 kg y MODR.
RESUMEN
Se determinó la concentración molar de los derivados de purinas (DP), creatinina (CT) en una muestra de orina de 63 vacas distribuidas en 4 grupos con diferentes niveles de alimentación (6.04, 10.09, 11.63 y 18.31 kg de MS y 2.98, 5.9, 7.37 y 12.5 kg MODR). A partir de la tasa DP/CT se calcularon las ecuaciones de regresión con el consumo de materia seca (MSI) y de materia orgánica degradable en el rumen (MODR). Además, se calculó la excreción diaria de los derivados de purinas (DPe) y con ella se obtuvo el rendimiento de nitrógeno microbiano por kilogramo de alimento ingerido (NM/kg).
La tasa molar DT/CT de una muestra de orina permitió discriminar entre los grupos de alimentación que difirieron en 4.5 kg de MSI o en 2.9 kg de MODR. Las ecuaciones de regresión encontradas fueron: DP/CT = 0.25 + 0.072 MSI + 0.002 MSI2 (r2 = 0.720) y DP/CT = 0.25 + 0.147 MODR + 0.0015 MODR2 (r2 = 0.710). Los valores estimados del total de los derivados de purinas excretados diariamente (DPe) permitieron discriminar entre niveles de alimentación de 1.54 kg de MS y de 1.47 kg de MODR, los valores encontrados por grupo fueron: I = 84.77 ± 12.19; II = 140.66 ± 7.21; III = 168.67 ± 6.45 y IV = 297.03 ± 9.72 (mmoles/d) (p < 0.05). Las ecuaciones de regresión entre los DPe y el consumo de alimentos fueron: DPe = 22.17 + 8.53 MSI + 0.364 X2 (r2 = 0.8) y DPe = 19.7 + 18.58 MODR + 0.281 X2 ( r2 = 0.798). El rendimiento calculado de NM/kg MS fue de 6.524 ± 0.928, 8.096 ± 0.549, 8.898 ± 0.491 y de 11.573 ± 0.74 g/kg siendo sólo distinto el grupo IV; mientras que expresados por kg de MODR el rendimiento fue estadísticamente igual para los 4 grupos, con valores de 13.224 ± 1.536, 13.846 ± 0.909, 14.041 ± 0.813 y de 16.953 ± 1.226 g/kg, respectivamente.
Se concluye que la tasa DP/CT de una muestra de orina de vacas presenta una alta correlación con el consumo de alimento y que los DPe estimados a partir de esta tasa permiten discriminar diferencias de consumo promedio de 1.5 kg de MS y de 1.47 kg de MODR.
AGRADECIMIENTOS
Los autores expresan su gratitud
al Dr. Mario Briones por su colaboración en el análisis estadístico,
al Prof. Sergio Recabarren por su traducción del resumen y al Dr.
René Garcés por su colaboración en la obtención
de las muestras de orina.
____________________________
Aceptado: 13.11.97.
* Financiado por Proyecto DIUC 93.153.001-4.
BIBLIOGRAFIA
ANTONIEWICZ, A.M., W. HEINEMANN, E.M. HANKS. 1981. Effect of level of feed intake and body mass on allantoin excretion and the allantoin to creatinine ratio in the urine of sheep, Rokz. Nauk Zoot. 8: 49-65.
ANTONIEWICZ, A.M., P.M. PISULEWSKI. 1982. Measurements of endogenous allantoin excretion in sheep urine, J. Agric. Sci. Camb. 98: 221-223.
ARAMBEL, M.J., E.E. BARTLEY, G.S. DUFVA, T.J. NAGARAJA, A.D. DAYTON. 1982. Effect of diet on amino and nucleic acids of rumen bacteria and protozoa, J. Dairy Sci. 65: 2095-2101.
ARC.1984. The nutrient requeriments of ruminant livestock (Suppl. 1), Commonwealth Agricultural Bureaux, Slough, U.K.
BALCELLS, J., J.A. GUADA, C. CASTRILLO, J. GASA. 1991. Urinary excretion of allantoin precursors by sheep after different rates of purine infusion into the duodenum, J. Agric. Sci. Camb. 116: 309-317.
BATES, D.B., J.A. GILLET, S.A. BARAO, W.G. BERGEN. 1985. The effect of specific growth rate and stage of growth on nucIeic acid-protein values of pure cultures and mixed ruminal bacteria, J An. Sci. 61: 713-724.
BRODY, S. 1945. Bioenergetics and growth. New York: Reinhold Publishing Corporation.
CHEN, X. B., F.D. DeB HOVELL, E.R. ORSKOV, D.S. BROWN. 1990a. Excretion of purines derivatives by ruminants: Effects of exogenous nucleic acid supply on purine derivative excretion by sheep, Br. J. Nutr. 63: 131-142.
CHEN, X.B., G. GRUBIC, E.R. ORSKOV, P. OSUJI. 1992a. Effect of feeding frequency on diurnal variation in plasma and urinary purine derivatives in steers, Anim. Prod. 55: 185-191.
CHEN, X.B., Y.K. CHEN, M.F. FRANKLIN, E.R. ORSKOV, W.J. SHAND. 1992b. The effect of feed intake and body weigth on purine derivative excretion and microbial protein supply in sheep, J. An. Sci. 70: 1534-1542.
CHEN, X.B., M.J. GOMES. 1992. Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives - an overview of the technical details. International Feed Resources Unit. Rowett Research Institute, Bucksburn, Aberdeen AB2 9SB, UK. Occasional Publication.
ERFLE, J.D., S. MAHADEVAN, F.D. SAUER. 1979. Effect of diet quality on adenosine 5'-triphosphate concentration and adenylate energy charge of rumen microbes from fistulated cows, J. Dairy Sci. 62: 248-291.
FAICHNEY, G.J., R.J. WELCH, G.H. BROWN. 1995. Prediction of the excretion of allantoin and total purine derivatives by sheep from the "creatinine coefficient", J. Agr. Sci. Camb. 125: 425-428.
FORBERG, C.W., K. LAMB. 1977. Use of adenosine-5'-triphosphate as an indicator of the microbiota biomass in rumen contens, Appl. and Env. Micr. 33: 528-537.
FUJIHARA, T., E.R. ORSKOV, P.J. REEDS, D.J. KYLE. 1987. The effect of protein infusion on urinary excretion of purine derivatives in ruminants nourished by intragastric nutrition, J. Agric. Sci. Camb. 109: 7-12.
GIESECKE, D., M. STANGASSINGER, W. TIEME-YER. 1984. Nucleic acid digestion and urinary purine metabolites in sheep nourished by intragastric infusions, Can. J. An. Sci. 64: 144-145. Suppl.
GONDA, H.L., J.E. LINDBERG. 1994. Evaluation of dietary nitrogen utilization in dairy cows based on urea concentrations in blood urine and milk, and on urinary concentration of purine derivatives, Acta Agr. Scan. Section A. An. Sc. 44 (4): 236-345.
LINDBERG, J. E. 1985. Urinary allantoin excretion and digestible organic matter intake in dairy goats Swedish, J. Agric. Res. 15: 31-37.
LINDBERG, J. E. 1989. Nitrogen metabolism and urinary excretion of purines in goat kids, Br. J. Nutr. 61: 39-321.
LINDBERG, J.E., K.G. JACOBSON. 1990. Nitrogen and purine metabolism at varying energy and protein supplies in sheep sustained on intragastric infusion, Br. J. Nutr. 64 (2): 359-370.
ORSKOV, E.R., F.D. DeB HOVELL, F.L. MOULD. 1980. The use of the nylon bag technique for the evaluation of feedstuff, Trop. Anim. Prod. 5: 195.
ORSKOV, R.E., N.A. MacLEOD. 1982a. The determination of the minimal nitrogen excretion in steers and dairy cows and its physiological and practical implications, Br. J. Nutr. 47: 625-636.
RESlNES, J. A., M.J. ARIN, M.T. DIEZ.1992. Determination of creatinine and purine derivatives in ruminant's urine by reversed-phase high-performance liquid chromatography, J. Chrom. 607: 199-202.
SUSMEL, P., B. STEFANON, E. PLAZZOTTA, M. SPANGHERO, C.R. MILLS. 1994. The effect of energy and protein intake on the excretion of purine derivatives, J. Agr. Sci. Camb. 123: 257-265.
SUSMEL, P., B. STEFANON, C.R. MILLS, M. COLITTI. 1991. Determinazione dell'adenosin-5'trifosfato in bioluminiscenza e suo impiego come indicatore della biomasa batterica nel liquido ruminale, Zootecnia e Nutrizione Animale, 17: 227-238.
SUSMEL, P., E. PLAZZOTTA, C.R. MILLS, B. STEFANON. 1993. Determination of RNA and ATP in the rumen liquid of cows fed with diets differing in forage to concentrate ratio, J. Sc. Food and Agr. 63: 39-45.
VERBIC, J., X.B. CHEN, N.A. MacLEOD, E.R. ORSKOV. 1990. Excretion of purine derivatives by ruminants. Effect of microbial nucleic acid infusion on purine derivative excretion by steers, J. Agr. Sci. Camb. 114: 243-248.
VERCOE, J.E. 1976. Urinary allantoin excretion and digestible dry-matter intake in cattle and buffalo, J. Agric. Sci. Camb. 86: 613-615.
WOLSTRUP, J., K. JENSEN. 1976. Adenosine triphosphate in the bovine rumen during maximum nutrient supply and starvation, J. Appl. Bact. 41: 243-250.