COMUNICACIONES
SUMMARY
Caseins are a family of milk proteins that exist in several molecular forms and are the main proteins present in the bovine milk. Genetic variants of these proteins have been associated with the quality and quantity of cheese derived from milk. Genotypes of 278 Frisón Negro Chileno cows were determined for k-casein by restriction fragment length polymorphism analysis of amplified DNA. A 350 bp fragment of the genomic bovine k-casein gene was amplified by PCR. Two HINF I sites were found in the amplified fragment of allele A, one at position 134 and one at position 266; only the latter site is present in allele B. Thus, digests of alleles A yielded 84 bp and 132/134 bp bands and digests of alleles B resulted in 84 bp and 266 bp bands. These bands, and thus genotypes AA, AB and BB, were recognised by agarose gel electrophoresis and ethidium-bromide staining. This technique was used to determine the k-casein allelic frequency in a Frisón Negro dairy herd.
The distribution of genotypes was slightly different, and the gene frequencies similar to those reported in the literature.
This molecular genetic technique based on molecular markers allows direct genotyping for milk k-casein with certainty and accuracy in bulls and females to be used in programs of dairy cattle improvement. Therefore, an early and precise identification of milk protein genotypes should have a direct impact on dairy cattle breeding strategies.
Palabras claves: k-caseína, PCR, genotipo.
Key words: k-casein, PCR, genotype.
INTRODUCCION
El mejoramiento de la calidad de la leche y sus propiedades de elaboración es una meta que desde hace muchos años encaran diferentes grupos de investigadores distribuidos en todo el mundo (Aschaffenburg, 1968; Grosclaude, 1988).
En Chile, el rubro leche juega un rol preponderante en el sector silvoagropecuario, estimándose que el 49.4% de la producción de leche de la IX y X Regiones se destina a producción de quesos y leche en polvo (ODEPA, 1997). Por lo tanto, para estas regiones el contenido de sólidos de la leche y particularmente la fracción proteica debiera tener prioridad dentro de los planes de mejoramiento bovino. Por otra parte, debido a los efectos perjudiciales que se asumen para la salud humana a consecuencia del alto consumo de grasas y colesterol, los cánones de consumo están marcando una clara tendencia a la disminución del contenido de grasas y a un aumento en el contenido y calidad proteica de la leche (Bradley, 1989; Hettinga, 1989). Actualmente, el 95% de las proteínas de la leche bovina han sido caracterizadas y clasificadas de acuerdo a sus propiedades bioquímicas. La caseína ha sido la más estudiada y es la proteína de mayor importancia para la industria lechera. Comprende cuatro principales familias de polipéptidos: as1, as2, b y k representando cerca del 80% del total de las proteínas de la leche (Stewart y col., 1984). El 20% restante corresponde a las dos principales proteínas del suero,a-lactoalbúmina y b-lactoglobulina. (Jenness, 1996). A su vez, cada una de estas proteínas posee diferentes variantes alélicas que son el resultado de la sustitución de unos pocos aminoácidos dentro de la secuencia proteica (Aschaffenburg, 1968). Tales variantes alélicas están estrechamente relacionadas con la calidad de la leche y especialmente con sus propiedades de elaboración. De esta forma, en el caso de la k-caseína, la variante alélica B está asociada con un incremento en el rendimiento de los quesos. Así, quesos producidos con este tipo de leche poseen un mayor contenido proteico, mayor rendimiento y mejor calidad que aquellos quesos producidos desde leches que poseen la variante alélica A. Tecnológicamente, en la elaboración de quesos, en base a leche con la variante alélica B de la k-caseína, se produce una cuajada más firme, hay un menor tiempo de coagulación y un mayor rendimiento bruto de queso.
Las diferentes variantes genéticas de caseína son controladas por genes autosómicos, los cuales son transmitidos desde los padres a la descendencia en forma mendeliana (Aschaffenburg, 1968). En el caso de la k-caseína, los bovinos heredan de cada progenitor un alelo A o B, conformándose así los genotipos homocigotos AA o BB, y el genotipo heterocigoto AB. Para poder seleccionar los toros en base a estos genotipos se utilizan pruebas de progenie muy extensas y que en general demandan entre 6 y 7 años (Medrano y Córdova, 1990). El prolongado período de tiempo requerido para tales propósitos hace poco práctico establecer programas de mejoramiento genético conducentes a incrementar la frecuencia génica de algún alelo deseado en la población. Aún más, la caracterización genotípica en las hembras se limita al período de lactancia, por lo que indefectiblemente debe esperarse a contar con animales adultos para poder estudiar los perfiles electroforéticos de las proteínas de la leche. (Schmidt, 1964). Con el advenimiento de tecnologías de punta, como la amplificación in vitro de ADN (PCR) "Polimerase Chain Reaction" (Mullis y Falloona, 1987; Saiki y col., 1988), actualmente es factible analizar directamente el genoma, lo que permite identificar en forma precisa las diferentes formas alélicas en el ganado lechero .
El objetivo del presente trabajo tiene por finalidad establecer en Chile las metodologías de trabajo que permitan la identificación de las formas alélicas prevalentes de la k-caseína para ser aplicadas al mejoramiento genético y para avanzar en la selección de caracteres de importancia económica en el ganado lechero. Junto con ello determinar la frecuencia genotípica y alélica de este gen en el rebaño de pedigree de Carillanca.
MATERIAL Y METODOS
a)Obtención de muestras. Se utilizaron muestras de sangre obtenidas de 278 ejemplares que conforman un rebaño de bovinos de la raza Frisón Negro Chileno, pertenecientes al Centro Regional de Investigación de Carillanca, distante 20 km de la ciudad de Temuco. Las muestras se obtuvieron de acuerdo a métodos convencionales de extracción de sangre (5 a 10 ml por animal) usando como anticoagulante EDTA-potásico 1.5 mg/ml de sangre.
b) Extracción de ADN: El aislamiento y purificación de ADN a partir de sangre total se realizó de acuerdo al método descrito por Weimberg (1991), el cual rinde ADN en cantidad y calidad suficientes para la reacción de polimerasa en cadena (PCR). Se utilizó además un método alternativo de extracción de ADN a partir de sangre total, el cual no requiere de extensivas etapas de purificación; es el procedimiento descrito por Singer-Sam y col. (1989), en el cual se aprovecha la capacidad quelante que tiene la resina Chelex-100 para extraer ADN desde células.
c) Amplificación de ADN in vitro. En base a la secuencia del gen de la k-caseína (Alexander y col., 1988) se diseñaron y sintetizaron dos oligonucleótidos o partidores de 25 y 26 mer respectivamente (Centro de Síntesis de Oligonucleótidos), adyacentes a los extremos de un fragmento de 350 pares de bases de dicho gen (Medrano y Córdoba, 1990). La reacción estándar para un volumen final de 30 µl contenía: 3 µl de tampón 10X de PCR (500 mM KCl, 100 mM Tris-HCl pH 8.3 y 25 mM MgCl2), 100 mM de cada dNTP, 0.75 U de Taq ADN polimerasa (Gibco BRL), 0.2 µM de cada partidor, alrededor de 30 ng de ADN y H2O desionizada estéril hasta completar volumen. Las muestras se amplificaron en un Termociclador Perkin-Elmer cuyo perfil térmico incluyó una fase de desnaturalización inicial a 94°C /5 minutos y 35 ciclos de 94°C/45 segundos, 65°C/60segundos, 72°C/30segundos para finalizar con una extensión final de 72°C/7 minutos.
d) Digestión del producto amplificado. Para diferenciar las variantes alélicas, el producto de la reacción de amplificación se digirió con HINF I. Para ello se utilizó la mitad del volumen de la reacción de PCR, al cual se le adicionaron 5 U de la enzima (Gibco BRL) y 10% del volumen final de reacción del tampón 10X React 2 de BRL (10 mM Tris-HCl pH 7.5, 10 mM MgCl2, 50 mM NaCl, 1 mM DTT). Cada reacción se incubó a 37°C por 90 minutos y se detuvo agregando 1 µl de tampón de carga (0,25% azul de bromofenol, 0.25% de xilen cyanol, 30% de glicerol y TAE 6X).
e) Determinación de las variantes alélicas. La identificación de las variantes alélicas se realizó por electroforesis en geles de agarosa al 3% en tampón TAE (Tris-acetato 0.04 M pH 8.0 y EDTA 1 mM). La corrida electroforética se realizó durante 120 minutos a un voltaje de 2.5 V/cm. Finalmente los geles se tiñeron con bromuro de etidio 0.5 mg/ml y se visualizaron en un transiluminador UV con lámpara de 312 nm.
f) Determinación de las frecuencias genotípicas y alélicas. En atención a que la determinación de las variantes alélicas permitió conocer el genotipo de cada animal muestreado, las frecuencias genotípicas y alélicas se determinaron por el método del recuento simple. Además se analizó el rebaño respecto a la condición de equilibrio génico según Hardy-Weinberg (Hoenigsberg, 1992).
RESULTADOS Y DISCUSION
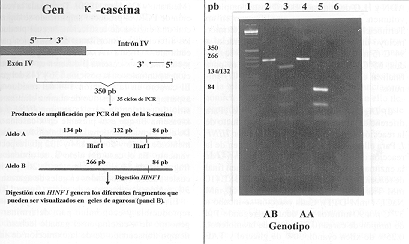
Las variantes alélicas se identificaron mediante la amplificación de un fragmento de ADN de 350 pares de bases que corresponde al gen de la k-caseína (Alexander y col., 1988). Este fragmento incluye 201 pares de bases del exón IV y 149 pares de bases del intrón del gen de la k-caseína (Medrano y Córdova, 1990). Terminada la reacción de PCR, el producto amplificado se digirió con una enzima de restricción, la cual produce los diferentes polimorfismos de restricción para cada alelo (figura 1A). Las variantes alélicas, k-caseína A y k-caseína B difieren en dos mutaciones puntuales en la posición 136 y 148 del gen. El cambio en la posición 148 del residuo de asparragina por el residuo de alanina elimina un sitio de restricción para la enzima HINF I en k-caseína B, que está presente en k-caseína A. De esta forma se obtiene para el alelo A un fragmento de 84 pb y dos de 134 y 132 pb, respectivamente. En el caso del alelo B, se obtiene un fragmento de 84 pb y uno mayor de 266 pb, los cuales fueron visualizados en geles de agarosa al 3%, teñidos con bromuro de etidio (figura 1B).
El procedimiento utilizado resultó ser preciso, reproducible y económico para determinar el genotipo de k-caseína en el ganado lechero. El tiempo transcurrido desde la extracción de sangre hasta la identificación del genotipo en el gel de agarosa ofrece enormes ventajas, comparado con el tiempo que se necesita para evaluar a un animal por métodos convencionales. El método demostró ser aplicable tanto a bovinos con relación de parentesco inmediato, independiente de la edad y etapa de lactancia, como también a animales de diferentes sexos (resultados no mostrados). El procedimiento de PCR utilizado permitió establecer la frecuencia alélica para cada uno de los alelos de la k-caseína, en un rebaño de bovinos de alta producción de leche. Como se observa en el cuadro 1, las frecuencias genotípicas obtenidas corresponden a un 65.8% para el homocigoto AA, 33.1%, para el heterocigoto AB y de un 1.1% para el homocigoto BB. Al comparar los resultados con estudios realizados en California (Medrano y Córdova, 1990) y Dinamarca (Beck y Kristiansen, 1990) (cuadro 1), se observa que las frecuencias genotípicas son similares, pero diferentes para los tres genotipos posibles respecto de los valores de Chile. En cuanto a la frecuencia alélica, en el rebaño Carillanca el alelo A representa un 82.4% y el B 17.6%. Estos valores son similares a los de los estudios realizados en California y Dinamarca por los autores mencionados. Como puede observarse, poblaciones razas similares ubicadas distantes unas de otras no se diferencian en la frecuencia alélica del gen de la k-caseína. En el cuadro 2 se muestran las frecuencias genotípicas observadas y las esperadas bajo la condición de equilibrio génico. En este caso las frecuencias de D, H, R (AA, AB, BB) no son iguales ap2 2pq q2. En consecuencia, la población no se encuentra en equilibrio y no existe cruzamiento al azar para este locus, lo que es consistente con el hecho de haber usado algunos toros heterocigotos para el gen de la k-caseína.
Figura 1. a) Representación esquemática del gen
de la k-caseína y sitios de reconocimiento
para la enzima de restricción HINF I. b) Electroforesis de
agarosa al 3% de los alelos de la k-caseína
de dos hembras. Carril 1 corresponde al estándar de tamaño
molecular 1 Kb 0.5 mg/µl (Gibco BRL), carriles 2 y 4 corresponden
al producto de amplificación de cada animal sin digerir, carriles
3 y 5 corresponden a los productos de amplificación digeridos con
HINF I indicando la presencia de un ejemplar heterocigoto AB (carril
3) y de un ejemplar homocigoto AA (carril 5) y carril 6 corresponde al
control negativo sin ADN.
a) Schematic representation of the k-casein
gen and HINF I restriction sites. b) 3% agarose gel separation of
k-casein alleles from two females. Lane 1 shows
a standard of molecular size 1 Kb 0.5 mg/ml (Gibco BRL) marker, lanes 2
and 4 carry an amplified segment of each animal without digest, lanes 3
and 5 represent an amplified segment corresponding to DNA genotypes AB
(lane 3) and AA (lane 5), and lane 6 is a negative control without DNA.
El procedimiento de amplificación por PCR constituye una poderosa herramienta para el análisis genético de las diferentes formas alélicas presentes para un determinado gen. Asimismo, esta herramienta puede ser usada para el diagnóstico de enfermedades genéticas en el ganado. Como el ADN puede ser extraído desde tipo celular (incluyendo espermatozoides), el método descrito aquí puede ser aplicado en programas de mejoramiento genético que utilicen inseminación artificial. De esta forma se podrán dirigir las estrategias de inseminación a fin de incorporar el alelo deseado en la población, o bien restringir la incorporación de aquellos genes deletéreos, determinando para ello la presencia y/o ausencia de estos genes en los progenitores. Esta forma de selección genética de bovinos, asistida por marcadores moleculares aplicada en toros, junto con la determinación de las diferentes formas alélicas presentes en las hembras, nos permitirá hacer una selección genética por marcadores moleculares de k-caseína certera y precoz de los progenitores a usar en los diferentes programas de mejoramiento genético. Finalmente, el uso de estas metodologías permitirá avanzar más rápido en el cambio genético de la población de vacas lecheras, posibilitando la selección de animales para estos y otros marcadores genéticos asociados a caracteres de importancia económica.
Comparación de la frecuencia genotípica y alélica para el locus de la k-caseína en estudios realizadosen USA, Dinamarca y Chile.
| Raza |
|
|||
| AA | AB | BB | Nº animales | |
| Holstein (USA) | 81.3% | 15.6% | 3.1% | 32 |
| Frisón Negro Danés | 73.5% | 22.9% | 3.6% | 223 |
| Frisón Negro Chileno | 65.8% | 33.1% | 1.1% | 278 |
|
Raza
|
|
|
|
|
|
|
| Holstein (USA) | 89.1% | 10.9% |
| Frisón Negro Danés | 85.0% | 15.0% |
| Frisón Negro Chileno | 82.4% | 17.6% |
|
|
||||
| AA | AB | BB | Nº animales | |
| Observada
Esperada |
183.0
188.6 | 92.0
80.7 | 3.0
8.6 | 278
278 |
| Proporciones | Total | |||
| Observada
Esperada = p2 2pq q2 p = q = |
0.658
0.678 0.824 | 0.330
0.290 | 0.010
0.031
0.176 | 1
1 |
RESUMEN
La caseína es la principal proteína presente en la leche de bovinos. Está constituida por una familia de proteínas, que a su vez existen en varias formas moleculares. Variantes genéticas de estas proteínas se han asociado a la calidad y cantidad del queso obtenido con este tipo de leche.
El objetivo del presente estudio tuvo por finalidad establecer las metodologías de trabajo que permitan la identificación de las formas alélicas prevalentes de la k-caseína para ser aplicadas al mejoramiento genético y avanzar en la selección de caracteres de importancia económica en el ganado lechero. Junto con ello, determinar la frecuencia genotípica y alélica de este gen en el rebaño de pedigree de Carillanca.
En este trabajo se utilizó el procedimiento de amplificación por PCR para determinar la frecuencia alélica del gen de la k-caseína bovina en un rebaño de bovinos Frisón Negro Chileno. Se amplificó por PCR un fragmento de 350 pb del gen bovino de la ?-caseína. El alelo A presenta 2 sitios de reconocimiento para la enzima de restricción HINF I, uno en la posición 134 y un segundo sitio en la posición 266. El alelo B presenta sólo este último. Así, la digestión con HINF I del alelo A origina bandas de 84, 132 y 134 pb respectivamente y la digestión del alelo B, produce bandas de 84 y 266 pb respectivamente.
La distribución de los diferentes genotipos en esta raza lechera varió
levemente con respecto a los estudios realizados en Dinamarca y EE.UU., sin
embargo, la frecuencia alélica de cada gen no se diferencia mayormente.
___________________________
Aceptado: 02.06.98.
BIBLIOGRAFIA
ALEXANDER, L.J., A.F. STEWART, A.G. MACKINLAY, T.V. KAPELINSKAYA, T.M. TKACH,S.I. GORODETSKY. 1988. Isolation and characterization of the bovine k-casein gen, Eur. J. Biochem. 178: 395-401.
ASCHAFENBURG, R. 1968. Genetic variants of milk proteins: their breed distribution, J. Dairy Res. 35: 447-459.
BECH, A.M. R.K. KRISTIANSEN. 1990. Milk protein polymorphism in Danish dairy cattle and the influence of genetic variants on milk yield, J. Dairy Res. 57: 53-62.
BRADLEY, R. L. Jr. 1989. Removal of cholesterol from milk fat using supercritical carbon dioxide, J. Dairy Sci. 72: 2834-2840.
GROSCLAUDE, F. 1988. Le polymorphisme genetique des principales lactoproteines bovines, Prod. Anim. 1: 5-17.
HETTINGA, D. H. 1989. Why alter milk composition?, J. Dairy Sci. 72: 2790-2800.
HOENIGSBERG, F.H. 1992. Genética de poblaciones. De Geminis Ltda., Santa Fe de Bogotá.
JENNESS, R. 1986. Biochemical and nutritional aspects of milk and calostrum. En : Larson, B.L. (ed.), Lactation. Iowa State University Press, Ames, Iowa.
MEDRANO, J. F., E. A. Cordova. 1990. Genotyping of bovine kappa-casein loci following DNA sequence amplification. Bio/technology 8: 144-146.
MULLIS, K. B., J. FALLOONA. 1987. Specific synthesis of DNA in vitro via polimerase catalized chain reaction. In "Methods in Enzymology". (Wu, R. (ed.), Vol. 155: 335-350.
ODEPA. 1997. Industria Láctea : Avances de recepción y producción. Chile. Ministerio de Agricultura. Oficina de Estudios y Políticas Agrarias, Dpto. de Información Agraria. 17 pp.
SAIKI, R. K., D. H. GELFAND, S.STOFFEL, S. J. SHARF, R. HIGUCHI, G. T. HORN, K. B. MULLIS, H. A. HERLICH. 1988. Prime directed enzymatic amplification of DNA with a thermostable DNA polymerase, Science 239: 487-491.
SCHMIDT, D. G. 1964. Starch gel electrophoresis of k-casein, Biochem. Biophys. Acta. 90: 411-414.
STEWART, A. F., Y. M. WILLIS, A. G. MACKINLAY. 1984. Nucleotide sequences of bovine as1- and k-casein cDNAs, Nucleic Acids Res. 12: 3895-3907.
SINGER-SAM, J., R. TANGUAY, A. RIGGS. 1989. Use of chelex-100 to improve the PCR signal from a small number of cells. Amplifications 3: 11.
WEIMBERG, G. 1991. A rapid method for preparing DNA from blood, suited for PCR screening of transgenes in mice, PCR Methods Appl. 1: 72-74.