COMUNICACIONES
Modificación del epitelio de revestimiento del útero y
desarrollo trofoblástico a los 21, 23, 28 y 36 días de la gestación en la vaca
Uterine lining modification and trophoblastic
development in the cow, at days 21, 23, 28 and 36 of gestation
DLAIKAN, H., M.V.; HERNANDEZ, A., M.V., M.Sc., Ph.D.; CORTES,
A., M.V.
Universidad Nacional de Colombia. Facultad de Medicina Veterinaria
y de Zootecnia. Bogotá. Colombia.
SUMMARY
In order to evaluate possible morphological modifications during implantation
in the cow, histological studies of the uterine lining epithelium and the
trophoblast were carried out in 8 heifers (2 per gestational stage) at
21, 23, 28 and 36 days of gestation. Ten samples were taken from each uterus,
and stained for light microscopical evaluation. Embryo location was assessed
by direct observation on days 23, 28 and 36 of pregnancy.
All embryos were located caudal to the mid uterine region. The more
relevant changes were: 1) modification of the uterine lining epithelium
(from a columnar or seudostratified columnar epithelium, to a cuboidal
and later to a flattened simple epithelium with sincitia). 2) The development
of the trophoblast (change from a simple cuboidal epithelium to a two-layered
cuboidal or columnar one). These modifications are initiated near the embryo
to spread peripherally afterwards, but they do not appear to be the same
in all regions of the uterus or the conceptus. Likewise, they are not seen
in a similar extent in conceptuses of the same age. A developed trophoblast
is typical of a viable conceptus, but a more precise change of implantation
is the presence of a flattened uterine lining epithelium. Implantation
in the cow occurs similar to sheep, although somewhat slower. The modulation
of trophoblast development could be due genetic programming and/or to paracrine
or autocrine secretions such as growth factors.
Key words: cow, implantation, uterus, trophoblast, placenta.
Palabras claves: vaca, implantación, útero, trofoblasto,
placenta.
INTRODUCCION
La mortalidad embrionaria afecta considerablemente la eficiencia reproductiva
en vacunos. Una parte importante de la pérdida de embriones sucede
durante la implantación (Brijaldo y col.,
1982; Sreenan y Diskin, 1986).
Aún no está claro si existen períodos de mayor
o menor vulnerabilidad del conceptus, ni se conocen completamente las características
anatómicas que permitan diferenciar los embriones viables de los
de dudosa viabilidad.
En la oveja, la implantación es un fenómeno lento y gradual,
en comparación con lo que sucede en especies (por ejemplo primates
y roedores) en las cuales hay un contacto íntimo entre el trofoblasto
y los tejidos maternos (Hernández, 1971;
Gaviria y Hernández, 1994; Guillomot,
1995). La estructura histológica del trofoblasto cambia entre
los 14 y los 17 días de gestación, de un epitelio simple
cúbico a uno biseriado cuboidal. Al comienzo de la implantación,
el epitelio de revestimiento del útero se modifica; pasa de cilíndrico
simple o seudoestratificado (típico de lo observado durante el ciclo
estral) a un tejido simple cúbico y posteriormente simple plano
(Gaviria y Hernández, 1994).
Existen evidencias (en observaciones hechas en especímenes de
edad gestacional superior a los 27 días) de que en la vaca podría
darse un tipo de implantación con características similares
a las observadas en la oveja (Hernández,
1975; Jiménez y Hernández, 1982).
Sin embargo, no aparecen en la literatura estudios cronológicos
acerca de los posibles cambios histológicos del trofoblasto y epitelio
uterino durante la implantación temprana en la vaca.
El presente trabajo tuvo como objetivo describir el desarrollo trofoblástico
y la modificación del epitelio uterino a los 21, 23, 28 y 36 días
de gestación en la vaca.
MATERIAL Y METODOS
Se utilizaron 8 hembras bovinas de la raza Holstein Friesian, entre
los 17 y los 20 meses de edad, dos por cada una de las siguientes edades
de gestación: 21, 23, 28 y 36 días.
Los animales fueron mantenidos en semiestabulación, bajo condiciones
adecuadas de nutrición (pasto cortado, suplementación con
concentrado comercial y sal y agua a voluntad) y manejo. Los signos de
estro fueron detectados con una hembra androgenizada provista con un chaleco
marcador, además de la observación de cada animal, tres veces
diarias. Se realizó monta directa, con un macho de la raza Normando.
El diagnóstico de la gestación se llevó a cabo
por observación del retorno o no al estro y mediante el cálculo
de los niveles de progesterona en el suero. Para esto último, se
tomaron muestras de sangre de la vena yugular, los días 5, 10, 15,
17, 19 y 21 (día 0= presentación de calor) para las gestaciones
de 21 días. Para las de 23, el mismo esquema, añadiendo el
día del sacrificio. Para las de 28 días, se tomaron muestras
en los mismos días usados para las gestaciones de 23 días.
Para las preñeces de 36 días, se tomaron muestras en los
mismos días de las gestaciones de 28 días, añadiendo
una toma el día 25.
Los animales fueron sacrificados, mediante sangría por sección
de las venas yugulares y carótidas, previa insensibilización.
Inmediatamente después del sacrificio, se removó el tracto genital,
el cual fue fijado con glutaraldehído tamponado al 2%. A continuación,
se hizo postfijación por inmersón con solución tamponada
de formalina al 10%. En cada cuerno uterino se establecieron 5 puntos equidistantes,
comenzando desde la extremidad útero-tubárica hasta el cuerpo
del útero. Así, el punto más cercano a la unión
útero-tubárica se identificó como el número 1 y
el más próximo al cuerpo del útero, como el 5. En cada
punto de divisón se realizaron cortes transversales, de los cuales se
obtuvieron, para colorear con Hematoxilina y Eosina, secciones de 5 micrómetros
de grosor previamente incluídas en parafina, mediante procedimientos
estándar. Posteriormente se analizó, en toda el área ocupada
por cada corte, el porcentaje de desarrollo trofoblástico (T) y el de
la modificación del epitelio uterino (MEU). Para este efecto, se midó
la longitud del corte y en base a ella se establecó que porcentaje de
la misma estaba ocupada por trofoblasto desarrollado o por un epitelio uterino
modificado (cúbico o cilíndrico).
Lo anterior se llevó a cabo utilizando un equipo computarizado
para análisis de imágenes (LECO 2000, USA), utilizando la
función de medición directa, debidamente calibrada para uso
con el objetivo de 40X. El epitelio corónico de dos o más
capas celulares fue considerado como un trofoblasto desarrollado. El útero
de un bovino no gestante con actividad ovárica normal está
recubierto de un epitelio que varía entre cilíndrico simple
y seudoestratificado (Priedkalns, 1976),
de tal manera que la presencia de un epitelio de otro tipo, fue tomado
como modificación.
Además se determinó el sitio preferencial de ubicación
de los embriones mediante observación directa, en las preñeces
de 23, 28 y 36 días. A los resultados obtenidos se les aplicaron
cálculos de estadística descriptiva.
RESULTADOS Y DISCUSION
En todas las edades de gestación estudiadas, T fue mayor en los cortes
del cuerno gestante al cuerpo lúteo que en los del no gestante. También
hubo variación en cuanto al T entre casos de la misma edad. Con el avance
de la gestación, T aumenta (ver figuras 1 y 2).
Sin embargo, en uno de los casos de 28 días de gestación, el valor
de T fue inferior al obtenido en todas las otras edades gestacionales (cuadro
1).
La MEU se hace de manera gradual. Inicialmente, el epitelio de revestimiento
del útero, seudoestratificado o simple cilíndrico, similar al
de animales no gestantes (figura 2), comienza a transformarse
en algunos sitios, en un epitelio de tipo cúbico (cuadro
4). Posteriormente, cambia a un tejido simple plano, con formación
de sincitios en algunas zonas (cuadro 5, figura
1). El epitelio de revestimiento del útero modificado, siempre se
observó en contacto con un trofoblasto desarrollado (figura
1). Los cambios se inician en el cuerno gestante al cuerpo lúteo,
predomi-nantemente en las zonas centrales y posteriores del mismo y luego se
extienden al cuerno no gestante. Existen variaciones individuales en relación
con MEU. A medida que avanza la gestación, puede observarse un aumento
en MEU, pues el porcentaje de epitelio plano es mayor a los 36 días que
a las demás edades estudiadas (cuadro 2).
|
|
 |
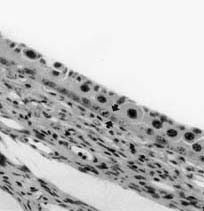
|
Figura 1. Trofoblasto desarrollado en contacto con el
epitelio de revestimiento endometrial modificado
(entre las flechas) en una zona del placentoma en la
vaca, a los 28 días de gestación. Aumento original
150X.
Developed trophoblast in contact with a modified lining
uterine epithelium (arrows) in a placentome. Twenty eigh days of gestation
in the cow. (Original magnification 150X).
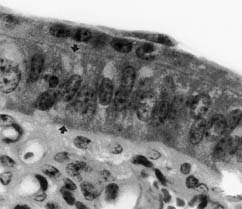
|
Figura 2. Trofoblasto no desarrollado en contacto con
el epitelio de revestimiento endometrial sin
modificación (entre las flechas) en una zona del
placentoma en la vaca, a los 28 días de gestación.
Aumento original 1000X.
Non-developed trophoblast in contact with an unmodified lining
uterine epithelium (arrows) in a placentome. Twenty eight days of gestation
in the cow. (Original magnification 1000X).
|
|
CUADRO 1. Valores promedio del porcentaje de desarrollo
trofoblástico del bovino en 4 edades de gestación.
Mean values of percentage of bovine trophoblastic development at 4
different stages of gestation
|
|
| Caso Nº |
Edad de Gestación (días) |
|
|
|
| 4 |
21 |
45 ± |
32.7 |
7.8 ± |
12.4 |
| 5 |
21 |
53.7 ± |
28.3 |
33.5 ± |
27.6 |
| 8 |
23 |
95.2 ± |
6 |
43.6 ± |
41.9 |
| 11 |
23 |
51.3 ± |
37.7 |
26.6 ± |
28.4 |
| 1 |
28 |
20 ± |
40 |
1.6 ± |
2.8 |
| 10 |
28 |
80 ± |
40 |
0.0 ± |
0.0 |
| 2 |
36 |
60 ± |
48.9 |
25.4 ± |
35.2 |
| 3 |
36 |
100.0 ± |
0.0 |
56.6 ± |
46.6 |
|
CG = Cuerno gestante; CNG = cuerno no gestante
* = Promedio ± desviación estándar
de T (datos obtenidos en todos los cortes observados).
|
|
CUADRO 2. Valores promedio del porcentaje
de dos tipos de epitelio revestimiento del útero bovino, en 4 edades
de gestación.
Mean values of the percentage of two types of uterine lining
epithelium in the bovine endometrium at 4 different stages of gestation.
|
|
| |
|
Epitelio cúbico
|
Epitelio plano
|
| Caso Nº |
Edad de gestación |
CG*
|
CNG*
|
CG*
|
CNG*
|
|
| 4 |
21 días |
43.1 ± |
34.4 |
8.5 ± |
14 |
0 ± |
0 |
0 ± |
0 |
| 5 |
21 días |
57.7 ± |
25 |
29.9 ± |
30.2 |
2.1 ± |
3.1 |
0 ± |
0 |
| 8 |
23 días |
74 ± |
29 |
43.6 ± |
37.4 |
19.8 ± |
33.5 |
0 ± |
0 |
| 11 |
23 días |
51.4 ± |
37.3 |
16.6 ± |
28.8 |
0 ± |
0 |
0 ± |
0 |
| 1 |
28 días |
16.6 ± |
31.7 |
3.0 ± |
4.4 |
19.2 ± |
38.4 |
0 ± |
0 |
| 10 |
28 días |
56.0 ± |
44 |
0.0 ± |
0.0 |
24 ± |
35.7 |
0 ± |
0 |
| 2 |
36 días |
8.5 ± |
15.7 |
36.0 ± |
37.2 |
51.5 ± |
44.4 |
0.6 ± |
1.2 |
| 3 |
36 días |
8.0 ± |
11.6 |
60.0 ± |
48.9 |
92.0 ± |
11.6 |
0 ± |
0 |
|
|
CG = cuerno gestante, CNG = cuerno no gestante,
* = promedio ± desviación estándar.
|
Los mayores valores de T se encuentran en las zonas posteriores del cuerno
uterino gestante (cuadro 3; cortes I3, I4 e I5), con excepción
del caso 4 de 21 días de edad, donde el mayor valor de T se obtuvo en
el corte más cercano a la unión útero-tubárica.
|
CUADRO 3. Porcentaje de desarrollo trofoblástico
en diferentes zonas del conceptus bovino en 4 edades de la gestación.
Percentage of trophoblastic development in different zones
of the bovine conceptus at 4 different stages of gestation.
|
|
| |
Cuerno gestante **
|
Cuerno no gestante
** |
| Caso |
EG* |
I1 |
I2 |
I3 |
I4 |
I5 |
C1 |
C2 |
C3 |
C4 |
C5 |
|
| 4 |
21 |
92 |
60 |
56 |
17 |
0 |
0 |
0 |
32 |
7 |
0 |
| 5 |
21 |
15 |
58 |
100 |
36 |
60 |
5 |
73 |
20 |
60 |
10 |
| 8 |
23 |
86 |
90 |
100 |
100e |
100e |
0 |
0 |
90 |
75 |
33 |
| 11 |
23 |
0 |
13 |
75 |
73 |
96e |
0 |
0 |
0 |
10 |
73 |
| 1 |
28 |
0 |
0 |
0 |
100e |
0e |
0 |
0 |
0 |
5 |
0 |
| 10 |
28 |
0 |
100 |
100 |
100e |
100e |
0 |
0 |
0 |
0 |
0 |
| 2 |
36 |
0 |
0 |
100e |
100e |
100 |
0 |
0 |
7 |
27 |
93 |
| 3 |
36 |
100 |
100 |
100 |
100e |
100 |
0 |
0 |
100 |
100 |
83 |
|
|
* EG= edad gestacional (en días). e = corte
cerca al cual estaba ubicado el embrión. ** I1 y C1 corresponden
a los cortes de las zonas más cercanas a la unión útero-tubárica;
I5 y C5, a los de las regiones más próximas al cuerpo del
útero.
|
|
CUADRO 4. Porcentaje de epitelio de revestimiento,
de tipo simple cúbico, en diferentes zonas del útero bovino,
en 4 edades de gestación.
Percentage of cuboidal uterine lining epithelium in different
zones in the bovine uterus, at 4 different stages of gestation.
|
|
| |
Cuerno gestante **
|
Cuerno no gestante
** |
| Caso |
EG* |
I1 |
I2 |
I3 |
I4 |
I5 |
C1 |
C2 |
C3 |
C4 |
C5 |
|
| 4 |
21 |
92 |
60 |
56 |
8 |
0 |
0 |
0 |
36 |
7 |
0 |
| 5 |
21 |
15 |
58 |
96 |
60 |
60 |
5 |
73 |
2 |
60 |
10 |
| 8 |
23 |
100 |
100 |
60 |
87e |
10e |
0 |
0 |
90 |
75 |
53 |
| 11 |
23 |
0 |
13 |
80 |
78 |
88e |
0 |
0 |
0 |
10 |
74 |
| 1 |
28 |
0 |
80 |
0 |
3e |
0e |
0 |
0 |
10 |
5 |
0 |
| 10 |
28 |
0 |
88 |
90 |
98e |
5e |
0 |
0 |
0 |
0 |
0 |
| 2 |
36 |
0 |
0 |
40e |
3e |
0 |
0 |
0 |
70 |
20 |
90 |
| 3 |
36 |
30 |
0 |
10 |
0e |
0 |
0 |
0 |
100 |
100 |
100 |
|
|
* EG= edad gestacional (en días). e = corte
cerca al cual estaba ubicado el embrión. ** I1 y C1 corresponden
a los cortes de las zonas más cercanas a la unión útero-tubárica;
I5 y C5, a los de las regiones más próximas al cuerpo del
útero.
|
|
CUADRO 5. Porcentaje de epitelio de revestimiento,
de tipo simple plano, en diferentes zonas del útero bovino, en
4 edades de gestación.
Percentage of flattened uterine lining epithelium in different
zones in the bovine uterus, at 4 different stages of gestation
|
|
| |
Cuerno gestante **
|
Cuerno no gestante
** |
| Caso |
EG* |
I1 |
I2 |
I3 |
I4 |
I5 |
C1 |
C2 |
C3 |
C4 |
C5 |
|
| 4 |
21 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| 5 |
21 |
0 |
0 |
8 |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
| 8 |
23 |
0 |
0 |
0 |
13e |
86e |
0 |
0 |
0 |
0 |
0 |
| 11 |
23 |
0 |
0 |
0 |
0 |
0e |
0 |
0 |
0 |
0 |
0 |
| 1 |
28 |
0 |
0 |
0 |
96e |
0e |
0 |
0 |
0 |
0 |
0 |
| 10 |
28 |
0 |
13 |
10 |
3e |
95e |
0 |
0 |
0 |
0 |
0 |
| 2 |
36 |
0 |
0 |
60e |
98e |
100 |
0 |
0 |
0 |
0 |
3 |
| 3 |
36 |
70 |
100 |
90 |
100e |
100 |
0 |
0 |
0 |
0 |
0 |
|
|
* EG= edad gestacional (en días). e = corte
cerca al cual estaba ubicado el embrión. ** I1 y C1 corresponden
a los cortes de las zonas más cercanas a la unión útero-tubárica;
I5 y C5, a los de las regiones más próximas al cuerpo del
útero.
|
De acuerdo con los resultados obtenidos, se puede inferir que el proceso de
la implantación en la vaca, es similar al correspondiente en la oveja,
iniciándose en las zonas cercanas al embrión para luego extenderse
a las áreas periféricas del cuerno gestante y luego al no gestante.
En la oveja, en el lapso de 10 días (entre los 14 y 24 de la gestación)
en preñeces únicas, el epitelio uterino ha sido modificado casi
en su totalidad (Gaviria y Hernández, 1994).
En el período de 17 días que cubre el presente estudio, la modificación
aludida no se produjo totalmente, lo cual estaría indicando un proceso
de implantación más lento en la vaca que en la oveja.
La amplia variación en cuanto al grado de desarrollo del trofoblasto
y de la modificación del epitelio uterino observada, puede implicar
la presencia de importantes diferencias, que podrían constituir
marcadores genéticos válidos de mayor o menor supervivencia
de los embriones. O por otro lado, la existencia de microambientes uterinos
más o menos propicios para el desarrollo del conceptus y
el éxito de la implantación.
El desarrollo del epitelio trofoblástico (paso de epitelio simple cúbico
a biseriado cúbico o cilíndrico) parece ser un requisito para
que se inicie la modificación del epitelio uterino. Esto puede depender
de programaciones genéticas del conceptus, o de las secreciones
del útero o el mismo trofoblasto. Hay varios estudios que indican que
el útero secreta los factores de crecimiento insulínico 1 y 2
(Geisert y col., 1991; Simmen
y col., 1993) y el factor epidérmico de crecimiento (Gharib-Hamrouche
y col., 1993), los cuales podrían influir en el desarrollo del conceptus.
Ello implicaría que dichos factores se elaboran y/o actúan de
manera preferencial en las cercanías del embrión. Sin embargo,
el desarrollo trofoblástico, no parece ser el único requisito
para que haya modificación del epitelio uterino, pues aún cuando
el grado de desarrollo del trofoblasto sea alto, no siempre corresponde a un
elevado porcentaje de epitelio modificado, como se observó en el presente
estudio a los 23 (caso 8) y a los 28 (caso 10) días de gestación.
Un trofoblasto conformado por un epitelio simple cúbico o aplanado,
podría ser tomado como característico de conceptus de dudosa
viabilidad (Boshier, 1968). Sin embargo, de acuerdo
con los hallazgos del presente estudio y lo encontrado previamente en la oveja
(Hernández, 1971; Gaviria
y Hernández, 1994), según el sitio de muestreo, aun en conceptus
viables se puede encontrar normalmente un trofoblasto como el descrito. Por
lo tanto, en las evaluaciones postmortem de viabilidad de los conceptus,
este aspecto debe tenerse en cuenta. Asimismo, según lo observado, el
desarrollo del trofoblasto parece ser un requisito indispensable para que se
den los cambios iniciales propios de la implantación (verbigracia modificación
del epitelio uterino). Por ende, el desarrollo del trofoblasto, o sea la presencia
de un trofoblasto conformado por epitelio cúbico o cilíndrico
biseriado, puede tomarse como un elemento más de evaluación de
la viabilidad del conceptus en bovinos, de igual manera como sucede en
la oveja (Gaviria y Hernández, 1994), en
adición al criterio histológico del número de células
binucleadas (Boshier, 1968) y al de la presencia
de vasos sanguíneos en las membranas embrionarias (Hernández,
1971). Sin embargo, el grado de modificación del epitelio de revestimiento
del endometrio, parece ser un cambio más confiable para evaluar el nivel
de éxito del proceso de la implantación, tanto en la vaca como
en la oveja. Esto puede ser de particular interés cuando no se disponga
de cortes de alantocorón, ya que a partir de cortes de tejido endometrial
podría evaluarse el grado de implantación y de acuerdo con la
extensón de los mismos, su expansón. Según los resultados
obtenidos en el presente estudio, hasta los 36 días de la gestación,
el cuerno no gestante, aún no ha participado en la implantación
del conceptus. Cabe anotar, en este contexto, que es posible que la porción
alantocorónica que ocupa el cuerno no gestante, haya sufrido el proceso
necrobótico, pues de acuerdo con un estudio previo, en la vaca se inicia,
aproximadamente a los 28 días de gestación (Hernández,
1975).
RESUMEN
Para evaluar las modificaciones morfológicas durante la implantación
en la vaca, se llevó a cabo un estudio histomorfométrico del epitelio
de revestimiento del útero y del trofoblasto en 8 novillas a los 21,
23, 28 y 36 días de gestación. Se utilizaron 10 cortes realizados
en puntos equidistantes de cada cuerno uterino. En las tres últimas edades
de gestación, se determinó la ubicación del embrión,
por observación directa.
Todos los embriones fueron encontrados en la mitad posterior del cuerno gestante.
Los cambios más relevantes encontrados se refieren a: 1) la modificación
del epitelio de revestimiento del útero (cambio de un epitelio cilíndrico
simple o seudoestratificado a uno cúbico simple y posteriormente a uno
de tipo plano simple con formación de sincitios), y 2) el desarrollo
trofoblástico (paso de un epitelio cúbico simple a uno biseriado
cúbico o cilíndrico). Ellos se hacen evidentes a medida que avanza
la gestación, pero no se llevan a cabo uniformemente en todas las regiones
del útero o del conceptus, ni suceden con dinámica similar
en los conceptus de la misma edad gestacional. Se inician en las cercanías
del embrión, para extenderse periféricamente. Un trofoblasto biseriado
cúbico o cilíndrico, es típico de un conceptus viable,
pero el cambio más significativo de la implantación, es la presencia
de un epitelio uterino de revestimiento modificado (simple plano con sincitios).
La implantación en la vaca es un proceso similar al informado previamente
en la oveja, aunque aparentemente más lento. La modulación de
los cambios sufridos por el trofoblasto, puede deberse a programaciones genéticas
del conceptus o a la secreción de factores de crecimiento secretados
por la madre y/o el conceptus.
AGRADECIMIENTOS
El presente trabajo fue auspiciado por la Universidad Nacional y Colciencias.
Los autores expresan su gratitud al Dr. Carlos Sánchez por su apoyo
en el procesamiento del material fotográfico.
Aceptado: 11.05.99.
BIBLIOGRAFIA
BOSHIER, D.P. 1968. Histological examination
of serosal membranes in studies of early embryonic mortality in the ewe,
J. Reprod. Fert. 15:81-86.
BRIJALDO, F., A. HERNÁNDEZ, H. SCHROEDER.
1982. Estudio de no retorno al calor en ganado Holstein Frisien de la Sabana
de Bogotá, Revista ACOVEZ 6:3-6.
GAVIRIA, M. T., A. HERNÁNDEZ. 1994.
Morphometry of implantation in the sheep. I. Trophoblast attachment, modification
of the.uterine lining, conceptus size and embryo location, Theriogenology.
41:1139-1149.
GEISERT, R.D., CH. LEE, A. SIMMEN, M.T. ZAVY,
A.F. FLISS, F.W. BAZER, R. SIMMEN. 1991. Expression of messenger RNAs encoding
insulin-like growth factor -I, II, and insulin-like growth factor protein-2
in bovine endometrium during the estrous cycle and early pregnancy, Biol.
Reprod. 45:975-983.
GHARIB-HAMROUCHE, N., N. CHENE,
M. GUILLOMOT, J. MARTAL. 1993. Localization and characterization of EGF/TGF-alpha
receptors on periimplantation trophoblast in sheep, J. Reprod. Fert.
98:385-392.
GUILLOMOT, M. 1995. Cellular interactions
during implantation in domestic ruminants, J. Reprod. Fert. Sup.
49:39-51.
HERNÁNDEZ, A. 1971. The development
of the extremities of the placenta of the domestic sheep. Thesis M.Sc.,
University of Bristol, England.
HERNÁNDEZ, A. 1975. Descripción
de las extremidades necróticas de la placenta de la vaca, ICA
10:235-238.
JIMÉNEZ, L., A. HERNÁNDEZ.
1982. Morfología del alantocorion bovino entre los 27 y 88 días
de gestación, Acovez 9:44-49.
PRIEDKALNS, J. 1976. Textbook of Veterinary
Histology. Lea & Febiger, Philadelphia.
SIMMEN, R. C. M., KO, F. A. SIMMEN, 1993.
Insulin-like growth factors and blastocyst development, Theriogenology
39:163-175.
SREENAN, J.M., M.G. DISKIN. 1986. The extent
and timing of embryonic mortality in the cow. En: J.M. Sreenan and M.G.
Diskin (eds.): Embryonic mortality in farm animals. A seminar of the CEC
programme of coordination of research on livestock productivity and management.
Martinus Nijhoff Publishers, Dordrecht, The Netherlands, pp. 1-7.