ARTÍCULOS ORIGINALES
S. E. RECABARREN, Biol., M.S.; A. LOBOS, MV, C. SCHNEIDER, MV.;
J. COX, MV.; J. A. PARILO, Ing Agr.
Laboratorio de Fisiología y Endocrinología Animal,
Facultad de Medicina Veterinaria, Universidad de Concepción,
Campus Chillán, CHILE.
SUMMARY
It has been proposed that blood-borne metabolic signals are captured by central or peripheral sensors to inform to the GnRH pulse generator about the metabolic status of the growing animal to make proper adjustments in the GnRH pulsatile secretion. During states of food restriction or fasting, the GnRH, and consequently, the LH secretion are reduced, probably due to inhibitory influences exerted by metabolic signals. Insulin has been suggested as a potential metabolic signal. However, little is known about the insulin sensitivity in prepubertal ewes, during experimental induction of negative energy balance, such as fasting, which may explain in part the effect of insulin as a metabolic signal. The purpose of this study was to establish the insulin sensitivity during fasting in ewe lambs. Six ewe lambs were given a bolus of insulin (0.1 IU/kg BW), one day before (day 0), on day 1, on day 8 of fasting and 24 hours (day 9) after the end of the 8-days fast. Plasma glucose concentrations were measured at 0,3,5,7,10, and 15 min after the insulin administrations to calculate the insulin sensitivity index (ISI= glucose at time 0 (G0) minus glucose at time 15, obtained from a regression curve, divided by G0). Plasma insulin concentrations were measured at-5, 0,3,5,7,10,15,20, and 60 min after insulin administration. On the other hand, plasma LH concentrations were measured at 0,10,20,40 and 60 min after insulin administrations in each day of the study to determine the effect of insulin on the acute LH secretion. Basal plasma concentrations of cortisol on day 0, day 1, 8 and 9 were also determined. The ISI did not change from day 0 to day 1, but decreased from day 1 to day 8 of fasting, range: 0.21 to 0.28 and -0.36 to 0.22 respectively with a median of 0.23 and 0.02 respectively (P<0.05). The ISI values on day 9 were not recovered; range: 0.02 to 0.28, with a median of 0.09. Plasma glucose concentrations descended linearly after insulin on day 0 and on day 1 of fasting, but not in day 8 and 9. Plasma insulin concentrations increased abruptly, reaching a peak at 3 min, and descending slowly thereafter in each day of the study. Plasma LH concentrations did not change with the insulin administrations. Fasting reduced basal plasma levels of glucose and LH and tended to reduce plasma insulin concentrations (P<0.08). Cortisol concentrations did not change during fasting. Results suggest that the insulin sensitivity is reduced with fasting in ewe lambs. A 24 hr refeeding is not sufficient to recover the insulin sensitivity, neither the LH basal plasma concentrations. This reduction in insulin sensitivity may allow other homeostatic mechanisms to operate to maintain plasma glucose levels. The insulin challenge did not affect plasma LH concentrations. Absence of changes in basal plasma cortisol concentrations suggests that fasting did not activate the adrenal axis.
Palabras claves: insulina, ayuno, borregas, LH, cortisol.
Key words: ewe lambs, LH, insulin, cortisol.
INTRODUCCION
Variada evidencia sugiere que durante estados de restricción alimenticia, tanto natural como experimental, la fertilidad de las hembras de los mamíferos disminuye, debido fundamentalmente a una disminución en la secreción de LH (Cameron y col., 1993). Se considera que la causa de la menor secreción de LH es una menor actividad del llamado centro hipotalámico generador de pulsos de GnRH, el cual mediante mecanismos aún no bien caracterizados, recibe información sobre el estatus metabólico del animal para finalmente readecuar la secreción de GnRH.
Se ha propuesto a la insulina como una posible señal metabólica que informaría al hipotálamo del estatus metabólico del animal (Steiner y col., 1983). De acuerdo a esta proposición, la disminución en la insulina plasmática sería reconocida por el hipotálamo, reduciendo la secreción de GnRH. Por el contrario, un aumento en la insulina estimularía la secreción de GnRH. Sin embargo, resultados contradictorios obtenidos sobre el efecto de la insulina en la secreción de LH, han hecho difícil la aceptación de esta hormona como señal metabólica que modula la secreción de GnRH (Hileman y col., 1993; Miller y col., 1995; Williams y col., 1996; Miller y col., 1998). Un aspecto que no se ha analizado en los estudios sobre la función de la insulina como señal metabólica es la sensibilidad tisular a la insulina en períodos de balance energético negativo, como es el ayuno.
El objetivo del presente trabajo fue caracterizar la sensibilidad tisular a la insulina mediante el cálculo del índice de sensibilidad a la insulina (ISI) antes, durante y después de un ayuno en ovejas prepúberes. Además, se determinó el efecto de la insulina sobre la secreción aguda de LH y las concentraciones plasmáticas basales de insulina, glucosa, LH y cortisol en respuesta al ayuno.
MATERIAL Y METODOS
Procedimientos generales. Se utilizaron 6 ovejas Suffolk down, de 16 semanas de edad, provenientes de la Unidad de Producción Ovina de la Facultad de Agronomía de la Universidad de Concepción, destetadas a los 3 meses de edad y mantenidas en pradera natural y suplementadas con alimento peletizado. Dos días antes de los experimentos, las borregas se trasladaron a la sala de experimentación, procediendo a cateterizar ambas venas yugulares bajo anestesia local con catéteres Mediziv G 18, de acuerdo a lo descrito anteriormente, (Recabarren y col., 1995). Las borregas se colocaron en bretes individuales con libre acceso al alimento peletizado y agua hasta el momento de los experimentos. Se procedió a realizar muestreos sanguíneos de acostum-bramiento el día anterior al inicio del estudio. El ayuno consistió en el retiro de la alimentación por un lapso de 8 días, permitiendo solo el brebaje, y se inició inmediatamente después de finalizado el muestreo sanguíneo del primer experimento de determinacion de la sensibilidad a la insulina (día 0). La realimentación se inició al término del muestreo sanguíneo correspondiente al día 8 de ayuno.
Determinación del índice de sensibilidad a la insulina (ISI). Antes del inicio del ayuno (día 0), el día 1, el día 8, y 24 horas después de terminado el ayuno (día 9) se procedió a realizar el estudio de la sensibilidad a la insulina. Para ello, se administró por medio de uno de los catéteres intrayugulares 0,1UI de insulina humana (Elli Lilly) por kilo de peso vivo, disueltas en 2 mL de suero salino. Se colectaron muestras de sangre con el catéter intrayugular contralateral (3 mL) a los 15, -5, 0, 3,5,7,10, 15 y 20 minutos post-administracion de insulina , las que se repartieron en tubos que contenían heparina y fluoruro al 3% para la medición de glucosa, y en tubos con heparina para la medición de hormonas. Se tomaron muestras adicionales a los 40 y 60 min para la medición de LH, insulina y cortisol.
El ISI se calculó de acuerdo a Grulet
y col. (1993) con la siguiente formula:
ISI=G0-G15/G0, donde G0 es la concentración plasmática de glucosa
al tiempo 3 min, y G15 es la concentración de glucosa al tiempo 15 min
determinada por el análisis de regresión de las concentraciones
plasmáticas de glucosa.
Medición de hormonas. LH. Las muestras de los tiempos 0,10,20,40 y 60 min se ocuparon para la medición de LH por radioimmunoensayo (RIA) de acuerdo a procedimientos descritos anteriormente (Recabarren y col, 1996). Todas las muestras se procesaron en un RIA. El coeficiente de variación intraensayo fue de 5%, el límite de sensibilidad fue de 0.1 ng/mL, definido como el 90% del buffer control. Las muestras del tiempo 0 sirvieron además para la medición de la LH basal de los días 0,1,8 y 9 del experimento.
Insulina. Las muestras de los tiempos 5,0,3,5,7,10,15,20 y 60 min se utilizaron para la medición de insulina por RIA (DPC, USA). El coeficiente intraensayo fue del 3%, con un nivel de sensibilidad de 5 uUI/mL, definido como el 90% del buffer control. Las muestras de los tiempos 5 y 0 se ocuparon en la medición de las concentraciones basales de insulina de los días 0,1,8 y 9.
Cortisol. Las muestras del tiempo 15 se ocuparon para la cuantificacion del cortisol basal, por RIA (DPC, USA). El coeficiente intraensayo fue del 6%, con un nivel de sensibilidad de 1 ug/dL, definido como el 90% del buffer control.
Medición de glucosa. La glucosa se cuantificó por el método de la glucosa oxidasa usando reactivos comerciales (Wiener Lab), con un coeficiente de variación intraensayo del 2%. Las muestras 5 y 0 sirvieron para determinar la glucosa basal de cada uno de los días del experimento.
Análisis estadístico. Las concentraciones plasmáticas de glucosa desde los 0 a los 15 min se analizaron por análisis de regresión simple para la determinación de la concentración de glucosa del minuto 15, necesaria para el cálculo del ISI. Además, las concentraciones de glucosa desde el tiempo 0 al tiempo 20 min se analizaron por análisis de varianza (ANDEVA) para muestras repetidas y comparación de los promedios con el test de Newman Keuls para determinar el efecto de la insulina sobre la glucosa plasmática. Las concentraciones de LH desde el tiempo 0 al tiempo 60 se analizaron con ANDEVA para muestras repetidas y comparación de los promedios con el test de Newman Keuls para determinar el efecto de la insulina sobre la secreción aguda de LH. Las concentraciones basales de LH, insulina, glucosa y de cortisol de los 4 días del experimento, se analizaron también con ANDEVA para muestras repetidas y comparación de los promedios con el test de Newman Keuls para definir el efecto del ayuno sobre las concentraciones basales de esos 4 parámetros. El ISI se analizó con el test no paramétrico de Kruskal Wallis. Se consideró para todos los análisis un P<0,05 como estadísticamente significativo. Los datos se presentan como promedio ± error estándar rango y mediana.
RESULTADOS
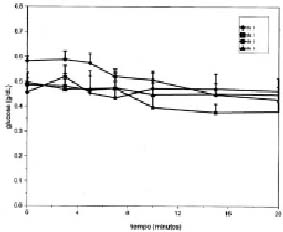
La administración endovenosa de insulina disminuyó significativamente la glucosa plasmática el día 0 (P<0,02) y el día 1 de ayuno (P<0,001). La primera caída significativa con respecto al tiempo 0 se produjo a los 10 min. Las concentraciones plasmáticas de glucosa disminuyeron levemente en respuesta a la insulina el día 8 de ayuno y 24 horas después de finalizado el ayuno (día 9) (figura 1). La diferencia en el efecto de la insulina sobre la glucosa plasmática, de acuerdo al día del ayuno, se relaciona con el ISI. El día 0, el ISI presentó un rango de 0,11 a 0,37 y el día 1, de 0,21 a 0,28, con una mediana de 0,23 y 0,23 respectivamente. Por el contrario, el ISI descendió significativamente el día 8, con un rango de 0,36 a 0,22 y una mediana de 0,02 (P<0,05), el cual no se modificó el día 9: rango de 0,02 a 0,28, y una mediana de 0,09.
La administración de insulina exógena aumentó significativamente las concentraciones plasmáticas de la hormona, con un máximo a los 3 min, en cada uno de los días del ensayo. Las concentraciones descendieron paulatinamente a partir de ese momento, hasta los 60 min. A los 60 min, las concentraciones de insulina no se diferenciaron de las concentraciones al tiempo 0 min.
Figura 1. Concentraciones plasmáticas de glucosa (g/L, promedio ± eem) en respuesta a un bolo de insulina (0,1 UI/kg PV) antes (día 0), durante el día 1 y 8 de ayuno y 24 horas despues de finalizado el ayuno (día 9), en ovejas prepúberes de 16 semanas de edad.
Plasma glucose concentrations (g/L, mean ± sem) in response to a bolus administration of insulin (0.1IU/Kg BW) before (day 0), during day 1 and day 8 of fasting and 24 hours after the end of fasting (day 9) in ewe lambs of 16 weeks of age.
La administración de insulina no modificó las concentraciones plasmáticas de LH el día 0, 1 y 8 de ayuno, mientras que el día 9, las concentraciones plasmáticas del tiempo 60 min fueron mayores que las del tiempo 0 (P<0,05).
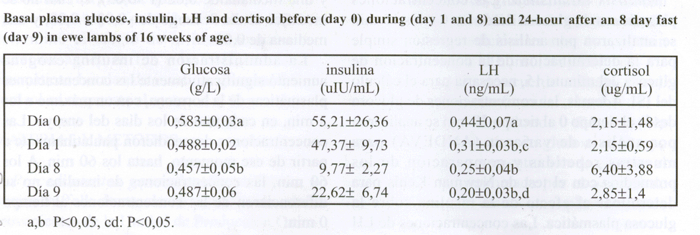
El cuadro 1 muestra las concentraciones basales de glucosa, insulina, LH y cortisol antes, durante y después del ayuno en ovejas prepúberes de 16 semanas de edad. Las concentraciones basales de glucosa disminuyeron desde el día 0 al día 8. Las concentraciones plasmáticas de LH descendieron con el ayuno, sin recuperarse con la realimentación (P<0,05), mientras que la insulina tendió a disminuir (P<0,08) desde el día 0 al día 8, aumentando ligeramente después de un día de realimentación (día 9). Las concentraciones plasmáticas basales de cortisol no se modificaron con el ayuno.
CUADRO 1. Concentraciones plasmáticas basales de glucosa, insulina, LH y cortisol antes (día 0), durante (días 1 y 8) y 24 horas (día 9) después de un ayuno de 8 días en ovejas prepúberes de 16 semanas de edad.
DISCUSION
Existen varios test que miden la sensibilidad tisular a la insulina. Uno de ellos es el llamado test de tolerancia a la insulina, el cual presenta una alta correlación con otros métodos más engorrosos, caros y con una alta ocupación en mano de obra (Grulet y col., 1993; Sir-Petermann y col., 1997). Los resultados del presente trabajo muestran que es un test altamente sensible para discriminar los efectos que tiene el ayuno sobre la sensibilidad tisular a la insulina. Los resultados muestran que la sensibilidad a la insulina disminuye drásticamente con el ayuno y que después de 24 horas de realimentación, la sensibilidad no se recupera totalmente. Esto significaría que durante el ayuno se desarrolla un estado de resistencia insulínica. La resistencia insulínica se define como el estado en el cual concentraciones normales de insulina producen una respuesta biológica atenuada (Krentz y Nattrass, 1996). Esto se corrobora con los efectos de la insulina sobre la glucosa plasmática en los días 8 y 9 del experimento, cuando la insulina exógena, es prácticamente ineficaz en reducir la glucosa plasmática. En ovejas se ha demostrado que durante la preñez (Petterson y col., 1993), y la lactancia (Vernon y col., 1990) se presenta resistencia insulínica, debido fundamentalmente a una reducción en la sensibilidad a la insulina en tejidos periféricos, principalmente tejido muscular y adiposo. La resistencia insulínica sería producto de alteraciones en el número de receptores a insulina o en defectos postreceptor (Sasaki, 1990). Durante el ayuno tendría como objetivo evitar el almacenamiento de glucosa en los tejidos sensibles a la insulina y para facilitar la gluconeogénesis (Yki-Jarvinen, 1993), y de esa forma mantener los niveles de glucosa necesarios para el metabolismo de los tejidos sensibles a las deficiencias de glucosa. Esta resistencia insulínica fisiológica sería un eficaz mecanismo que permitiría mantener la homeostasis de la glucosa plasmática en períodos críticos como es el ayuno. La homeostasis de la glucosa está bajo un control estricto entre el aporte y la utilización de la glucosa. La insulina juega un rol clave en este proceso inhibiendo la producción hepática y estimulando la captación de glucosa por los tejidos sensibles, en particular el tejido muscular esquelético y el tejido adiposo. La hipoinsulinemia y la resistencia que se origina durante el ayuno tendría, entonces, como objetivo facilitar las acciones fisiológicas de las hormonas contraregulatorias como ser glucagon, catecolaminas, cortisol y GH, con el fin de mantener los niveles plasmáticos de glucosa.
La hipótesis que postula a la insulina como señal metabólica sostiene que aumentos de insulina plasmática tendrían una acción estimulante sobre la secreción de GnRH/LH. La administración de insulina en el presente trabajo no tuvo efecto sobre la secreción aguda de LH. Estos resultados concuerdan con los de Hileman y col (1993), quienes al administrar insulina endovenosa e intracerebroventricular no observaron modificaciones en la secreción pulsátil de LH en ovejas prepúberes con alimentacion normal o con restricción alimenticia. Otros estudios en ovejas y ratas han mostrado una disminución en la secreción aguda de LH, pero en esos casos también podría atribuirse la reducción en la concentración plasmática de LH a la hipoglicemia que sigue a la administración de insulina (Clarke y col., 1990; Goubillon y Thalabard, 1996). Estos efectos de la insulina contrastan con los resultados encontrados en los estudios de Miller y col. (1995,1998), quienes demostraron que los aumentos de insulina plasmática con dietas ricas en lupino (que provee altas cantidades de energía digestible y proteína), se observan después de varios días y son paralelos en aumentos en la secreción de LH. Los resultados de Miller y col. (1995) concuerdan con los de Downing y col. (1995) y de Catalano y col. (1996), en los cuales la alimentación rica en energía aumenta los niveles de insulina y esta hormona se relaciona a su vez con la tasa ovulatoria. Es probable entonces que las acciones estimuladoras de la insulina sobre la secreción de GnRH/LH sean de largo plazo y producto de una hiperinsulinemia sostenida en el tiempo. Esta situación podría señalar más eficazmente el estado metabólico del animal a las neuronas GnRH, que el monitoreo de los cambios agudos de insulina plasmática.
Se ha postulado que el ayuno es un tipo de estrés, que al igual que con otros tipos de estrés, al activarse el eje adrenal, conduciría a un aumento en la liberación de CRH, ACTH y cortisol, repercutiendo en el eje hipotálamo-hipófisis-gónada como consecuencia de la inhibición de la secreción de GnRH por la CRH (Petraglia y col., 1987). En la rata existe evidencia que respalda esta hipótesis (Maeda y col., 1994), sin embargo, en el presente estudio así como en otros de nuestro laboratorio (Recabarren y col., 1999), las concentraciones basales de cortisol durante el ayuno no se modificaron, lo cual sugiere que el ayuno de ocho días no sería un estrés en la oveja prepúber y que el cortisol no estaría mediando los efectos del ayuno en la secreción de LH. Estos resultados concuerdan con los de L'Anson y col. (1994), quienes descartan que el cortisol influya en la disminución de la secreción de GnRH/LH durante períodos de restricción alimenticia en la oveja prepúber. Por otro lado, estudios en hombres y mujeres (Cameron y col., 1991; Olson y col., 1995) y monos (Schreihofer y col., 1993) reafirman la noción de que la restricción alimenticia y el ayuno no implican reacciones de estrés que inhiban la secreción de GnRH/LH. Estos resultados reafirman la hipotesis de que señales metabólicas serían más importantes que la activación del eje adrenal en la regulación de la secreción de GnRH/LH en estados de ayuno y probablemente en otros estados de balance energético negativo en la oveja prepúber.
En resumen, un ayuno de 8 días en ovejas prepúberes de 16 semanas de edad disminuye la sensibilidad tisular a la insulina. Las concentraciones basales de glucosa y LH, disminuyeron, las de insulina tendieron a disminuir; por el contrario, las concentraciones plasmáticas de cortisol no se modificaron con el ayuno. La administración de insulina no afectó la secreción de LH lo que sugirere que esta hormona no influye sobre la secreción aguda de LH, sin embargo, no se puede descartar que la dosis de insulina no sea suficiente, que la duración de la estimulación sea muy corta o que estén ausentes otras señales metabólicas que en forma concomitante influyan en la secreción de LH. El ayuno no activaría el eje adrenal, lo que sugiere que el ayuno no induciría estrés en la oveja prepúber.
RESUMEN
Con el fin de explicar las modificaciones en la secreción de LH que se observan durante el ayuno en animales, se ha postulado que señales metabólicas de origen sanguíneo serían captadas por sensores a nivel central o periférico, los cuales transmitirían la información sobre el estatus metabólico hacia las neuronas GnRH para producir ajustes en su secreción y consecuentemente en la de LH. Entre estas señales se ha postulado a la insulina. Es poco conocido el posible rol de la insulina como señal metabólica en ovejas prepúberes en estados de balance energético negativo experimental como por ejemplo el ayuno. La acción de la insulina en estas condiciones experimentales podría estar asociada a la sensibilidad tisular a la insulina. El objetivo del presente trabajo fue establecer la sensibilidad a la insulina en ovejas prepúberes de 16 semanas antes, durante y después de un ayuno corto de 8 días. Para ello, 6 ovejas Suffolk Down prepúberes recibieron un bolo de insulina (0,1 UI/kg PV) endovenosa el día anterior (día 0), el día 1 y el día 8 de ayuno y 24 horas después de finalizado el ayuno (día 9). Se midió glucosa plasmática a los 0,3,5,7,10, y 15 min después de la administración de insulina para calcular el índice de sensibilidad a la insulina (ISI= glucosa al tiempo 3 min (G0) menos la glucosa al tiempo 15 min, obtenida mediante el análisis de regresión de los valores de glucosa, dividido por G0). Se midió la concentración plasmática de insulina a los 5, 0,3,5,7,10,15,20, y 60 min después de la administración de insulina. Se midió también la LH plasmática a los 0,10,20,40 y 60 min después de la administración de insulina en cada uno de los días del ensayo para determinar el efecto de la insulina sobre la secreción aguda de LH. Las concentraciones basales de cortisol se midieron los días 0, 1, 8 y 9 del ensayo para reconocer el posible estrés inducido por el ayuno. EL ISI no cambió entre el día 0 y 1, pero disminuyó del día 1 al día 8 de ayuno con un rango de 0,21 a 0,28 y de -0,36 a 0,22 respectivamente, con una mediana de 0,23 y 0,02 respectivamente (P<0,05). Los valores de ISI el día 9 no se recuperaron a pesar de la realimentación, presentando un rango de 0,02 to 0,28, con una mediana de 0,09. Las concentraciones plasmáticas de glucosa descendieron con la administración de insulina el día 0 y 1 de ayuno pero no se modificaron el día 8 y 9. Las concentraciones de insulina aumentaron abruptamente alcanzando un valor máximo a los 3 min de su administración y descendiendo después. Las concentraciones plasmáticas de LH no se modificaron con la insulina exógena. El ayuno redujo los niveles basales de glucosa y de LH y tendió a disminuir los niveles basales de insulina (P<0,08). Las concentraciones basales de cortisol no se modificaron. Los resultados sugieren que la sensibilidad a la insulina disminuye con el ayuno y que la realimentación por 24 horas no permite la recuperación de ésta. La reducción en la sensibilidad a la insulina permitiría que otros mecanismos homeostáticos operen para mantener las concentraciones plasmáticas de glucosa. La ausencia de cambios en el cortisol plasmático sugiere que el ayuno no activaría el eje adrenal.
AGRADECIMIENTOS
Los autores expresan sus agradecientos a los Drs. Gordon E. Niswender y Leo E. Reichert Jr, y a la NIDDK de Estados Unidos por materiales para los radioimmunoensayos de LH. Al Dr. Marcelo Calvillán, del Laboratorio de Endocrinología y Metabolismo de la Facultad de Medicina Occidente de la Universidad de Chile por su colaboración en el cálculo del ISI.
_________________________________________
Aceptado: 08.08.2000.
* Este trabajo fue financiado por Proyecto DIUC 94.153.003-1.0.
BIBLIOGRAFIA
CAMERON, J.L., T. WELTZIN, C. McCONAHA, D.L. HELMREICH, W.H. KAYE. 1991. Suppression of reproductive axis activity in men undergoing a 48 hour fast. J. Clin. Endocrinol Metab. 73: 35-41.
CAMERON, J.L., D.L. HELMREICH, D.A. SCHREIHOFER. 1993. Modulation of reproductive hormone secretion by nutritional intake:stress signals versus metabolic signals. Hum Reprod 8 (suppl 2): 162-167.
CATALANO, R., L. SIRHAN, S. RECABARREN, B. MUÑOZ, H. MANTEROLA. 1995. "Flushing" en merino precoz.II. Efecto del nivel de energía y proteína sobre concentraciones plasmáticas de insulina, FSH y urea. Av. Prod Anim 20: 165-178.
CLARKE, I.J., R.J.E. HORTON, B.W. DOUGHTON. 1990. Investigation of the mechanism by which insulin-induced hypoglycemia decreases luteinizing hormone secretion in ovariectomized ewes. Endocrinology 127: 1470-1476.
DOWNING, J.A., J. JOSS, P. CONNEL, R.J. SCARAMUZZI. 1995. Ovulation rate and the concentrations of gonadotrophic and metabolic hormones in ewes feed lupin grain. J. Reprod Fert 103: 137-145.
GOUBILLON, M.L, J.C. THALABARD. 1996. Insulin-induced hypoglycemia decreases luteinizing hormone secretion in the castrated male rat: Involvement of opiate peptides. Neuroendocrinology 64: 49-56.
GRULET, H, V. DURLACH, A.C. HECART, A. GROSS, M. LEUTENEGGER. 1993. Study of the rate of early glucose disappearance following insulin injection: insulin sensitivity index. Diab. Res .Clin. Pract 20: 201-207.
HILEMAN, S.M, K.K. SCHILLO, J.B. HALL. 1993. Effects of acute, intracerebroventricular administration of insulin on serum concentrations of luteinizing hormone, insulin and glucose in ovariectomized lambs during restricted and adlibitum feed intake. Biol. Reprod. 48: 117-124.
KRENTZ, A.J., M. NATTRASS. 1996. Insulin resistance: a multifaceted metabolic syndrome. Insights gained using a low-dose insulin infusion technique. Diabet. Med. 13: 30-39
L'ANSON, H., E.H. QUINT, I.R. WOOD, B.G. ENGLAND, D.L. FOSTER. 1994. Adrenal axis and hypogonadotropism in the growth-restricted female lamb. Biol. Reprod. 50: 137-143.
MAEDA, K.I., F.R.A. CAGAMPANG, C.W. COEN, H. TSUKAMURA. 1994. Involvement of the catecholaminergic input to the paraventricular nucleus and of corticotropin-releasing hormone in the fasting-induced suppression of luteinizing hormone release in female rats. Endocrinology 134: 1718-1722.
MILLER, D.W., D. BLACHE, G.B. MARTIN 1995. The role of intracerebral insulin in the effect of nutrition on gonadotrophin secretion in mature male sheep. J. Endocrinol. 147:321-329
MILLER, D.W., D. BLACHE, R. BOUKHLIQ, J.D. CURLEWIS, G.B. MARTIN. 1998. Central metabolic messengers and the effects of nutrition on gonadotrophin secretion in sheep. J. Reprod. Fert. 112: 347:356.
OLSON, B.R., T. CARTLEDGE, N. SEBRING, R. DEFENSOR, L. NIEMAN. 1995. Short-term fasting affect luteinizing hormone secretory dinamics but not reproductive function in normal-weight sedentary women. J. Clin. Endocrinol. Metab. 80: 1187-1193.
PETRAGLIA, F., S. SUTTON, W. VALE, P. PLOTSKY.1987. Corticotropin-releasing factor decreases plasma luteinizing hormone levels in female rats by inhibiting gonadotropin-releasing hormone release into hypophysial-portal circulation. Endocrinology 120: 1083-1088.
PETTERSON, J.A., F.R. DUNSHEA, R.A. EHRHARDT, A.W. BELL. 1993. Pregnancy and undernutrition alter glucose metabolic responses to insulin in sheep. J. Nutr. 123: 1286-1295
RECABARREN, S.E. A. URRUCELQUI, M. ROBBIANO, A. LOBOS, P. ORELLANA, J.PARILO. 1995. Efecto de la infusión endovenosa de L-arginina y L-ornitina sobre la secreción de hormona del crecimiento en ovejas prepúberes. Arch. Med.Vet. 27: 99-104.
RECABARREN, S.E, H. ESCOBAR, A. LOBOS, M.P. RECABARREN, J. PARILO. 1996. Luteinizing hormone pulse frequency is increased by arginine infusion in prepubertal sheep. Exp. Clin. Endo. Diabetes 104: 74-79.
RECABARREN, S.E., A. LOBOS, E. ABALOS, C. ARRIAGADA.1999. Effect of naloxone infusions on the pulsatile LH secretion, insulin, and cortisol plasma concentrations in fasted ewe lambs. Exp. Clin. Endocrinol Diabetes 107: suppl. 1, S-48. Abstract P012. 43rd Symposium of the German Society of Endocrinology, Kiel, March 10-13, 1999.
SASAKI, S. 1990. Mechanism of insulin resistance in the post receptor events in sheep: 3-O-methylglucose transport in ovine adipocites. Horm. Metab. Res. 22: 457-461.
SIR-PETERMANN, T., T. CASTILLO, M. CALVILLAN, S. MUÑOZ, G. LOPEZ. 1997. Evaluación de la sensibilidad tisular a la insulina en mujeres normales, obesas y obesas hiperandrogénicas. Rev. Méd. Chile. 125: 977-985.
STEINER, R.A., J.L. CAMERON, T.H. McNEILL, D.K. CLIFTON, W.J. BREMNER. 1983. Metabolic signals for the onset of puberty. En: Neuroendocrine aspects of reproduction. R.L. Norman (ed.) Pp. 183-227, Academic Press, Nueva York.
VERNON, R.G., A. FAULKNER, W.W. HAY Jr., D.T. CALVERT, D.J. FLINT. 1990. Insulin resistance of hindlimb tissues in vivo in lactating sheep. Biochem. J. 270: 783-786
YKI-JARVINEN, H. 1993. Action of insulin on glucose metabolism in vivo. Bailliers Clin Endocrinol Metab. 7: 903-927.