ARTÍCULOS ORIGINALES
N. TADICH1,M.V., Ph.D.; C. GALLO2, M.V., Ph.D., M; ALVARADO1,M.V.
1 Instituto de Ciencias Clínicas
Veterinarias. 2 Instituto de Ciencia y Tecnología
de Carnes,
Facultad de Ciencias Veterinarias, Universidad Austral de Chile, Casilla 567,
Valdivia, Chile.
SUMMARY
An experiment was carried out in winter (June - July) with the aim of determining the effects of transporting cattle for up to 36 hours with and without a resting period, on the blood concentrations of cortisol, glucose, ß-HBA, PCV values and blood CK activity.
Forty A. Angus and Hereford cross steers and heifers from the same farm with milk teeth or two teeth and a mean live weight of 400 kg were randomly divided into two groups of 20 animals each. They were transported during 36 hours with and without a resting period of eight hours after the first 24 h of journey. The animals were slaughtered following a lairage period of 12 h after arrival to the abattoir.
Blood samples were obtained from the jugular vein at the farm on their arrival to the resting station on arrival to the abattoir and finally at slaughter when the animals were bled. The blood cortisol concentrations were determined by radioimmunoassay (RIA); the glucose blood concentrations by the GOD PAP test without deprotenization (GL 2623, RANDOX); the ß-HBA by using the enzymatic technique that uses the ß-hidroxibutirate deshidrogenase enzyme for measuring the transformation from NAD+ to NADH; the PCV values by the microhematocrite technique and the CK blood activity was measured by the UV- kinetic method at 340 nm and 37 C. Descriptive statistics were used for analysing the results.To determine the differences between means, ANOVA was used and Kruskal-Wallis when the variances were not homogenous.
A high individual variation in response to stress for transport was observed. It can be concluded that transport for 36 h with or without a resting period was detrimental for the welfare of the animals. This is supported by the increase on the blood concentrations of cortisol, glucose and CK activity from the farm, to arrival at the abattoir and stunning. However, the resting period had a beneficial effect on the blood activity of CK and PCV values but to a lesser extent on fat mobilisation as reflected by the blood concentrations of ß-HBA at stunning.
Palabras claves: estrés, transporte, bovinos.
Key words: stress, transport, cattle.
INTRODUCCION
En Chile existen 4.098.438 cabezas de ganado bovino, de las cuales 1.587.557 se encuentran en la Xa Región (38,7%), 784.336 en la IXa Región (19,1%) y sólo 164.014 en la Región Metropolitana (4,0 %) (Chile, 1997 a). De 1.094.684 cabezas beneficiadas en 1997, sólo 147.554 se beneficiaron en la Xa Región (13,5%) y 108.919 en la IXa Región (9,9%), mientras que 500.259 cabezas (45,7%) fueron beneficiadas en la Región Metropolitana (Chile, 1997b).
En Chile es característico el traslado en pie de un gran número de animales desde los centros productores a los centros de faenamiento y consumo. Según Matic (1997) más del 50% de los bovinos beneficiados en la planta faenadora de Lo Valledor, son transportados en camión por distancias superiores a los 600 km. Gallo y col. (1995) en un estudio realizado con 6.000 animales transportados desde Osorno a Santiago, comprobaron que el tiempo de viaje promedio era de 24 horas, con ayunos que alcanzaban las 60 horas al incluir la espera en matadero previo al sacrificio. Bustos (1997) señala un tiempo de transporte promedio de 20 horas y de espera en matadero (ayuno) superior a 24 horas, para novillos trasladados desde Valdivia a Santiago.
El transporte de animales en Chile, se rige por la Ley de Carnes o Ley 19.162 (Chile, 1992), con su Reglamento general de transporte de ganado y carne bovina (Chile, 1993), que establece los requisitos mínimos para los vehículos que transportan ganado, la vigilancia regular de los animales por el transportista durante el viaje y períodos de descanso con abrevaje por un mínimo de ocho horas, cada 24 horas de viaje continuo. Dicho reglamento establece que los animales deberán permanecer en espera, en la planta faenadora, por un lapso mínimo de 12 horas y máximo de 72 horas. La espera mínima no considera diferencias en relación al tiempo de transporte previo, ni tampoco hace diferencias entre especies animales. Según Gallo y col. (1995) los tiempos más frecuentes de espera en matadero están alrededor de las 36 horas.
El transporte y el ayuno producen diversos grados de estrés, según la duración e intensidad del estímulo, desencadenando diversas respuestas fisiológicas y conductuales adaptativas en los animales (Forrest y col., 1979; Lister y col., 1981; Mitchell y col., 1988; Shaw y Tume, 1992; Tume y Shaw, 1992; Warriss y col., 1995). El estrés que provocan los manejos a que son sometidos los animales antemortem, tiene un efecto directo sobre el bienestar animal e indirecto sobre la calidad y cantidad de carne producida (Warriss, 1992). Entre los diversos factores que inducen estrés en los animales se han señalado: la privación de alimento, las variaciones de temperatura, el transporte por períodos prolongados, el ejercicio muscular y los estímulos sociales, como es la mezcla de grupos de animales de distinto origen (Mitchell y col., 1988; Shaw y Tume, 1992; Tume y Shaw, 1992). También algunos de los factores implícitos en el transporte, tales como carga, descarga, arreo, hacinamiento, movimiento, ruido, vibraciones, y todos los manejos menores inmediatamente previos al sacrificio (Cockram y col., 1996; Gallo, 1994, 1996). Según Gallo (1996), en Chile se les da poca importancia a estos factores, a diferencia de lo que ocurre en países más desarrollados, en que se presta gran atención al bienestar animal.
El estrés en los animales se puede cuantificar mediante dos métodos: i) el análisis de la conducta animal; y, ii) mediciones en los tejidos y fluidos del animal (Shaw y Tume, 1992). En relación con este último punto, entre los cambios que se pueden medir en los animales, destacan la determinación de corticoesteroides, adrenalina, noradrenalina y hormonas tiroídeas. Otras variables asociadas al estrés, son los niveles sanguíneos de cortisol, glucosa, lactato, insulina, ácidos grasos volátiles y volumen globular aglomerado (VGA) (Forrest y col., 1979).
La mayoría de los investigadores utilizan la determinación de la concentración de cortisol plasmático como un indicador de estrés asociado con los manejos, transporte y sacrificio de los animales (Crookshank y col., 1979; Warriss y col., 1984; Warner y col., 1986; Shaw y Tume, 1992; Cooper y col., 1995; Horton y col., 1996). Estos mismos autores también han utilizado adicionalmente mediciones de volumen globular aglomerado (VGA), glucosa sanguínea y de enzimas como la creatinfosfoquinasa (CK). Warriss y col. (1995) señalan que en animales destinados al sacrificio también existe una influencia del comportamiento sobre la actividad plasmática de CK, concentración plasmática de ácidos grasos libres y betahidroxibutirato (ß-HBA), siendo los valores más altos en los animales sometidos a las condiciones más estresantes. Cooper y col. (1995) también han utilizado como indicadores de estrés en terneros la medición de progesterona y endorfinas.
No es de conocimiento de los autores que en Chile se hayan efectuado estudios sobre la influencia de tiempos de transporte terrestre prolongados sobre algunas variables indicadoras de estrés en bovinos. Tampoco ha sido posible encontrar en la literatura internacional estudios que utilicen tiempos de transporte de 36 horas. Por este motivo y considerando la reglamentación vigente, se consideró importante efectuar un estudio preliminar para determinar el efecto de un período de descanso sobre algunas variables indicadoras de estrés, en animales transportados vía terrestre durante 36 horas.
MATERIAL Y METODOS
El experimento se llevó a cabo durante los meses de junio y julio de 1998 en la provincia de Valdivia, Chile.
Material. Se utilizaron 20 novillitos y 20 vaquillas de similares características en cuanto a raza (A. Angus x Hereford), procedencia (San José de la Mariquina), edad (DL y 2D) y peso (400 kg en promedio). Para su transporte se utilizaron dos camiones que tenían la estructura indicada por la reglamentación actual (Chile, 1993), con características y capacidad similar. Se utilizó una estación de descanso ubicada en las cercanías de la ciudad de Valdivia.
Método. Los animales se dividieron al azar en dos tratamientos (T1 y T2) con 20 animales cada uno (10 hembras y 10 machos). En el predio de origen, posterior a la toma de muestras y previo al embarque, los animales fueron identificados con un arete plástico y pesados.
Tratamiento 1 (sin descanso): Este grupo fue transportado en forma continua, utilizando una carretera pavimentada durante 36 horas, sin alimentación ni descanso. La densidad animal en el camión era de 1.00 m2/500 kg de peso vivo.
Tratamiento 2 (con descanso): Este grupo fue transportado en condiciones de carretera durante 24 horas, desde el predio de origen hasta la estación de descanso; a su arribo fueron sangrados, pesados individualmente y luego permanecieron durante ocho horas recibiendo agua a discreción y pasto seco. Posterior al período de descanso fueron reembarcados en el mismo camión y transportados por otras 12 horas para completar el período de 36 horas de transporte. La densidad animal en el camión era de 1,04 m2/500 kg pv. Ambos grupos fueron descargados finalmente en la Planta Faenadora de Carnes (PFC) FRIVAL S.A., para ser faenados luego de aproximadamente 12 horas de reposo.
La extracción de las muestras de sangre en el predio el día del transporte, a la llegada a la estación de descanso y a la llegada a la PFC, se realizó por punción yugular. La muestra correspondiente al período de la sangría, correspondió a la muestra tomada inmediatamente comenzado el proceso de desangramiento del animal, posterior a su noqueo.
Análisis de las variables sanguíneas. Las variables determinadas fueron las concentraciones sanguíneas de cortisol, glucosa, betahidroxibutirato (ß-HBA), el hematocrito (VGA) y la actividad plasmática de la creatinfosfoquinasa (CK). Las muestras de sangre fueron obtenidas de acuerdo a los tiempos y grupos especificados anteriormente. Se utilizaron tubos con NaF para la determinación de la glucosa y tubos con heparina para la determinación de cortisol, VGA, ß-HBA y CK.
Determinación de la concentración plasmática de cortisol. Las concentraciones de cortisol plasmático se determinaron en duplicados de 20 ul mediante radioinmunoensayo (RIA), usando kits comerciales (DPC, USA), en la Facultad de Ciencias Veterinarias de la Universidad de Concepción.
Determinación del hematocrito: El VGA se determinó utilizando la técnica del microhematocrito descrita por Wittwer y Böhmwald (1983).
Determinación de la concentración plasmática de glucosa: Se realizó según el procedimiento de la prueba para glucosa GOD-PAP, sin deproteinización (RANDOX) utilizando un espectrofotómetro HITACHI 4020.
Determinación de la concentración plasmática de betahidroxibutirato: El plasma se sometió a una técnica enzimática para determinar la concentración sanguínea de ß-HBA, en la que el ß-HBA es oxidado por NAD+ (nicotinamida adenin dinucleótido), mediante la enzima 3 HBDH (3-hidroxibutirato deshidrogenasa) a acetoacetato. La cantidad de NAD+ reducida se midió en un espectrofotómetro HITACHI 4020 a 340 nm.
Determinación de la actividad plasmática de la creatinfosfoquinasa (EC 2.7.3.2): La actividad enzimática de la CK, se determinó mediante un método UV-cinético, a 340 nm y a 37C, optimizado según la DGKC. Se emplearon reactivos Boehringer Mannheim y un espectrofotómetro HITACHI 4020.
Análisis estadístico. Para el análisis de los resultados se utilizó estadística descriptiva, los que se presentan en forma de cuadros y gráficos, con promedios y desviaciones estándar. La significancia de la diferencia entre las medias de los distintos períodos en un mismo tratamiento y entre períodos para los distintos grupos, se determinó utilizando el análisis de varianza o ANOVA y el análisis de Kruskal-Wallis, cuando las varianzas no eran homogéneas. El programa utilizado fue el Epi Info versión 6,03 (Dean y col., 1997).
RESULTADOS
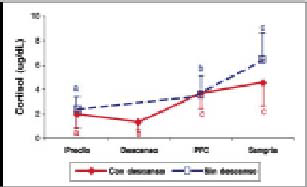
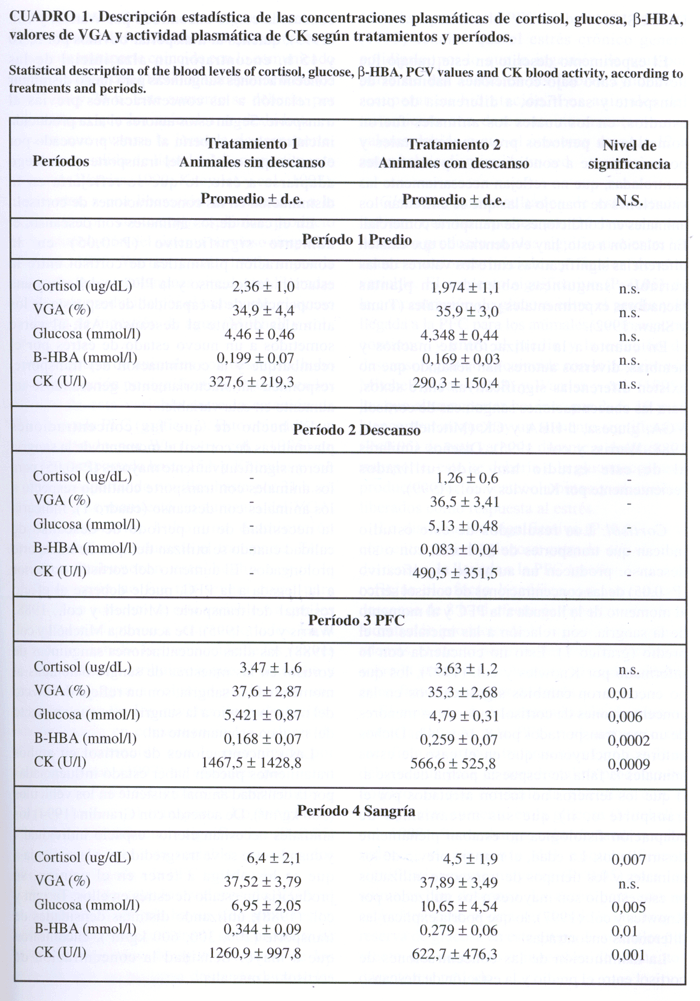
Cortisol. Las concentraciones plasmáticas del cortisol (gráfico 1) aumentaron en forma significativa (P<0,05) entre el predio y la PFC y el predio y la sangría en los dos tratamientos, excepto en los animales que tuvieron acceso a un período de descanso, en los cuales se observó una disminución estadísticamente significativa (P<0,05) entre los valores obtenidos en el predio y los obtenidos al llegar a la estación de descanso. En cuanto al análisis entre tratamientos sólo hubo diferencias estadísticamente significativas (P<0,05) al momento de la sangría (cuadro 1).
Valores de VGA. En ambos tratamientos se observó una tendencia no significativa (P>0,05) a aumentar en los valores de VGA entre el predio de origen y el momento de la sangría (gráfico 2). En los animales sin un período de descanso este aumento fue significativo (P< 0,05) entre el predio de origen y la PFC, manteniendo valores similares al momento de la sangría. En los animales con descanso los valores de VGA no tuvieron variaciones significativas (P>0,05) entre el predio de origen y la estación de descanso, sin embargo, tendieron a disminuir a la llegada a la PFC. En este mismo tratamiento se produjo un aumento significativo (P<0,05) de los valores de VGA entre la llegada a la PFC y el momento de la sangría. Al analizar las diferencias entre tratamientos para cada período sólo se observaron diferencias significativas (P<0,05) en los valores obtenidos a la llegada a la PFC (cuadro 1).
Concentraciones plasmáticas de glucosa. Se puede observar que hubo un aumento significativo (P<0,05) de las concentraciones plasmáticas de glucosa entre los distintos períodos para ambos tratamientos (gráfico 3). Al comparar las concentraciones plasmáticas de glucosa entre tratamientos, se encontraron diferencias significativas (P<0,05) a la llegada a la PFC y al momento de la sangría (cuadro 1).
Concentraciones plasmáticas de ß-HBA. Se observó una disminución significativa (P<0,05) en los valores obtenidos en el predio y el segundo muestreo en el grupo con descanso. Sin embargo, hubo un aumento significativo (P< 0,05) entre el predio y PFC para este tratamiento.
En ambos tratamientos hubo un aumento significativo (P<0,05) de las concentraciones plasmáticas de ß-HBA entre los valores iniciales y los valores obtenidos al momento de la sangría (gráfico 4). En este experimento se observaron diferencias significativas (P< 0,05) entre tratamientos a la llegada a la PFC y al momento de la sangría (cuadro 1).
Actividad plasmática de la CK. La actividad
plasmática de CK aumentó significativamente (P<0,05) entre
el predio de origen, la PFC y el momento de la sangría, en ambos grupos
(gráfico 5). Al comparar los tratamientos entre
sí, las diferencias significativas (P<0,05) se produjeron en las concentraciones
de la enzima a la llegada a la PFC y al momento de la sangría, siendo
más alta la actividad en el grupo sin descanso (cuadro
1).
Gráfico 1. Concentraciones plasmáticas de Cortisol (promedio ± d.e) en bovinos transportados con y sin descanso.
Plasmatic concentrations of Cortisol (mean ± s.d) in cattle transported by road with and without a rest.
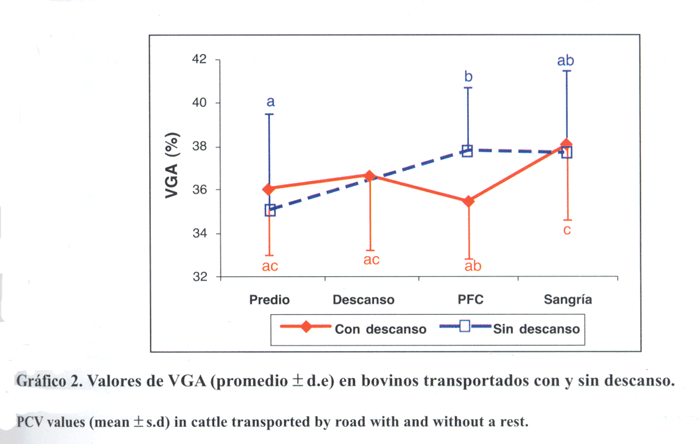
Gráfico 2. Valores de VGA (promedio ± d.e) en bovinos transportados con y sin descanso.
PCV values (mean ± s.d) in cattle transported by road with and without a rest.
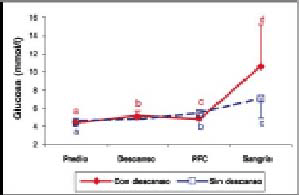
Gráfico 3. Concentraciones plasmáticas de Glucosa (promedio ± d.e) en bovinos transportados con y sin descanso.
Plasmatic concentrations of Glucose (mean (± s.d) in cattle transported by road with and without a rest.
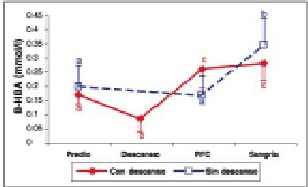
Gráfico 4. Concentraciones plasmáticas de ß-HBA (promedio ± d.e) en bovinos transportados con y sin descanso.
Plasmatic concentrations of ß-HBA (mean ± s.d) in cattle transported by road with and without a rest.
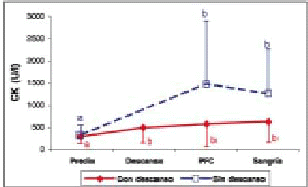
Gráfico 5. Concentraciones plasmáticas de CK (promedio ± d.e) en bovinos transportados con y sin descanso.
Plasmatic concentrations of CK (mean ± s.d) in cattle transported by road with and without a rest.
CUADRO 1. Descripción estadística de las concentraciones plasmáticas de cortisol, glucosa, b-HBA, valores de VGA y actividad plasmática de CK según tratamientos y períodos.
Statistical description of the blood levels of cortisol, glucose, b-HBA, PCV values and CK blood activity, according to treatments and periods.
DISCUSION
El experimento descrito en este trabajo fue llevado a cabo bajo condiciones habituales de transporte y sacrificio, a diferencia de otros estudios, en los cuales los animales fueron sometidos a períodos pre-experimentales y posteriormente a condiciones experimentales controladas, que no reflejan necesariamente las situaciones de manejo a las que se enfrentan los animales en condiciones de transporte comercial. En relación a esto, hay evidencias de que existen diferencias significativas entre los valores de las variables sanguíneas obtenidos en plantas faenadoras experimentales y comerciales (Tume y Shaw, 1992).
En cuanto a la utilización de machos y hembras, diversos autores han señalado que no existen diferencias significativas entre sexos, para las concentraciones sanguíneas de cortisol, VGA, glucosa, ß-HBA y CK (Mitchell y col., 1988; Warriss y col., 1995). Diseños similares al de este estudio han sido utilizados recientemente por Knowles y col. (1999).
Cortisol. Los resultados de este estudio indican que transportes de 36 horas, con o sin descanso, producen un aumento significativo (P<0,05) de las concentraciones de cortisol sérico al momento de la llegada a la PFC y al momento de la sangría, con relación a las iniciales en el predio (gráfico 1). Esto no concuerda con lo informado por Knowles y col. (1997), los que no encontraron cambios significativos en las concentraciones de cortisol en terneros menores de un mes transportados por 8, 16 y 24 h. Dichos autores concluyeron que en el caso de estos animales la falta de respuesta podría deberse a: i) que los terneros no fueron afectados por el transporte o, ii) que sus mecanismos de adaptación fisiológica no estaban plenamente desarrollados. La edad, el manejo previo de los animales y los tiempos de transporte utilizados en este estudio son mayores a los utilizados por Knowles y col. (1997), lo que podría explicar las diferencias encontradas.
La disminución de las concentraciones de cortisol entre el predio y la estación de descanso concuerda con lo encontrado por Warriss y col. (1995), quienes al transportar bovinos por 5, 10 y 15 h, encontraron un alza inicial de las concentraciones sanguíneas para luego disminuir en relación a las concentraciones previas al transporte. Según estos autores el alza producida inicialmente se debería al estrés provocado por el embarque e inicio del transporte, para luego adaptarse a éste, lo que se reflejaría en la disminución en las concentraciones de cortisol.
En el caso de los animales con descanso, el aumento significativo (P<0,05) en la concentración plasmática de cortisol entre la estación de descanso y la PFC se debería a una recuperación de la capacidad de respuesta de los animales durante el descanso. Así, al verse sometidos a un nuevo estado de estrés por el reembarque y la continuación del transporte, responden satisfactoriamente, generándose un aumento en esta variable.
El hecho de que las concentraciones plasmáticas de cortisol al momento de la sangría fueron significativamente mayores (P>0,05) para los animales con transporte continuo respecto a los animales con descanso (cuadro 1), indicaría la necesidad de un período de descanso de calidad cuando se utilizan tiempos de transporte prolongados. El aumento del cortisol, posterior a la llegada a la PFC, puede deberse al efecto residual del transporte (Mitchell y col., 1988; Warris y col., 1995). De acuerdo a Mitchell y col. (1988), las altas concentraciones sanguíneas de cortisol en las muestras de sangre obtenidas al momento de la sangría, son un reflejo del efecto del manejo previo a la sangría más que un efecto del noqueo propiamente tal.
Las concentraciones de cortisol en ambos tratamientos pueden haber estado influenciadas por la densidad animal existente en los vehículos (500 kg/m2). De acuerdo con Grandin (1994) los animales necesitan cierto "espacio individual", y cuando este se ve trasgredido por la cercanía a que se les obliga a tener en el camión, se produciría un estado de estrés en ellos. Tarrant y col. (1988), utilizando distintas densidades de transporte (200, 300, 600 kg/m2), encontraron que a mayor densidad la concentración de cortisol es más alta.
Hematocrito (VGA). La tendencia al aumento de los valores de VGA a medida que aumenta el tiempo de transporte coincide con lo comunicado por Knowles y col. (1997) al transportar terneros por 8, 16 y 24 h. Este aumento se debería, por una parte, a que los animales se deshidratan a medida que avanza la jornada de transporte, debido a la salida de líquido del compartimento vascular (Warriss, 1990; Knowles y col., 1997), y, por otra, a una liberación de eritrocitos a la circulación por contracción esplénica, producto de la estimulación del sistema nervioso simpático (Warriss, 1990; Warriss y col., 1995), o por la acción de las catecolaminas circulantes liberadas en el estrés producido por el transporte, el ayuno y la falta de ingestión de agua (Mitchell y col., 1988).
En el grupo de animales con descanso, hubo una disminución no significativa (P>0,05) en los valores de esta variable entre la estación de descanso y la PFC (gráfico 2). Esto coincide con lo encontrado por Warriss y col. (1995) en ganado transportado hasta 15 h y Knowles y col. (1999) transportando novillos hasta por 31 h, en los que no se manifestó un aumento del VGA, por el contrario, hubo una disminución progresiva de éste durante el transporte. Estos autores atribuyeron este hecho a que la deshidratación no fue tan severa como para producir un aumento de esta variable y a que los animales se adaptaron a ser transportados. En el caso de este estudio la disminución de los valores de VGA podrían atribuirse a que el descanso y el acceso al agua a voluntad producirían una recuperación de los niveles de hidratación, disminuyendo en consecuencia los valores de VGA. Esto coincide con lo encontrado por Knowles y col. (1997) en animales que fueron alimentados con soluciones de electrolitos y glucosa durante el transporte. Lo anterior se tradujo en una diferencia significativa (P<0,05) en los valores de esta variable entre los dos grupos a la llegada a la PFC (cuadro 1).
El hecho de que los valores de VGA al momento de la sangría hayan sido similares en ambos tratamientos (cuadro 1) sería producto de la mayor ingestión de agua por parte de los animales que no tuvieron descanso durante las 12 h de reposo en la PFC. Caballero y Sumano (1993) señalan que el estrés crónico genera polidipsia, por lo que la ingestión de agua por parte de los animales les habría ayudado a recuperar su hidratación. Los animales del tratamiento con descanso incrementaron sus valores debido a que el consumo de agua 12 h antes habría disminuido su sed.
Glucosa. Los resultados de este estudio indican que transportes por 36 h con o sin descanso producen un alza significativa entre las concentraciones iniciales y finales de esta variable. El aumento significativo (P<0,05) de la concentración de glucosa entre el predio y la llegada a la PFC para los animales con transporte continuo, y entre el predio y la estación de descanso, (gráfico 3) coincide con lo descrito por Mitchell y col. (1988) y Warriss y col. (1995), los que al transportar bovinos por 2, 5, 10 y 15 h encontraron que hay un aumento significativo (P<0,05) de esta variable durante y después del transporte, esto se debería a la gluconeogénesis producida por las catecolaminas y el cortisol liberados como respuesta al estrés.
La disminución significativa (P<0,05) de la concentración de glucosa entre la estación de descanso y la llegada a la PFC (gráfico 3) podría reflejar el efecto benéfico del período de descanso de 8 h, el efecto de recuperación de la hidratación producto de la ingestión de agua y, además, el efecto positivo del llenado del rumen sobre la sensación de hambre, el cual, también, es un factor generador de estrés.
El aumento de la concentración de glucosa entre la llegada a la PFC y el momento del sacrificio en ambos tratamientos (gráfico 3) se debería a un peak de noradrenalina producto del noqueo, lo que estimula la glicólisis con el consiguiente aumento en la glucosa (Mitchell y col. 1988). Sin embargo, cabe destacar que los valores al momento del noqueo fueron significativamente mayores (P<0,05) para los animales con descanso. La razón para esto no está clara, pero podría ser debido a una mayor reserva de glucosa en este grupo de animales por efecto del descanso e ingestión de alimentos durante ese período.
ß-hidroxibutirato (ß-HBA). Los resultados encontrados indican que transportes por 36 h, con o sin descanso, producen un alza significativa (P<0,05) entre los valores predio y los obtenidos al momento de la sangría para esta variable.
La disminución significativa (p< 0,05) de esta variable en las primeras 24 h de transporte concuerda con lo encontrado por Knowles y col. (1999) al transportar animales hasta por 31 h. Sin embargo, Warriss y col. (1995) y Knowles y col. (1997), al transportar terneros menores de un mes, por 8, 16 y 24 h y bovinos de 12 y 18 meses de edad por 5, 10 y 15 h, respectivamente, encontraron que al final del período más corto de cada experimento hubo una disminución de la concentración plasmática de ß-HBA y en los otros tiempos hubo un aumento en la concentración de este metabolito respecto al grupo control. Estos autores señalan que el alza en los valores de ß -HBA, al aumentar el tiempo de transporte, se debería a la movilización de las reservas grasas corporales, como respuesta a un ayuno cada vez más prolongado. En este estudio, la disminución observada en esta variable podría deberse a la utilización de concentraciones sanguíneas de ß-HBA circulante y el aumento posterior a una movilización de las reservas de grasas corporales y a la relación existente entre la gluconeogénesis y la producción de cuerpos cetónicos (Herdt, 1988).
De acuerdo a estos resultados y a los de Tadich y col. (1999), quienes utilizaron cuatro tiempos de transporte: 3, 6, 12 y 24 h, las concentraciones plasmáticas de ß-HBA no serían un buen indicador de estrés agudo, debido a la falta de sincronía entre el aumento de la glucosa y el de cuerpos cetónico, al menos durante las primeras 24 h de transporte
Creatinfosfoquinasa (CK). Los animales en ambos tratamientos tuvieron un alza significativa (P<0,05) en la actividad plasmática de CK entre el predio, la llegada a la PFC y el momento de la sangría (gráfico 5). Warriss y col. (1995) y Knowles y col. (1999) utilizando distintos tiempos de transporte encontraron que la actividad plasmática de CK aumentó al incrementarse el tiempo de transporte, indicando que éste es físicamente estresante. Esto concuerda con lo encontrado en este estudio y se debería al esfuerzo que realizan los animales para mantener la postura en un vehículo en movimiento, lo que les causa una gran fatiga muscular y en algunos casos contusiones; como resultado la CK es liberada producto de un cambio de permeabilidad en las membranas celulares y llega a la circulación desde el tejido muscular dañado, provocando el aumento en su actividad plasmática (Warriss, 1990; Warriss y col., 1995; Knowles y col., 1997; Knowles, 1999). Esto se vería confirmado por los resultados obtenidos por Gallo y col. (1998), los que encontraron un mayor número de contusiones en los animales transportados en forma continua durante 36 h.
La densidad de animales por m2 utilizada en este experimento (500 kg/m2) podría explicar, en ambos grupos, el aumento significativo (P<0,05) de los valores de CK al llegar a la PFC. La importancia de la densidad de transporte ha sido demostrada por Knowles y col. (1997), los cuales utilizando densidades de 129 y 122 kg/m2 no encontraron aumentos en la actividad plasmática de CK, debido a que los animales pudieron descansar durante el viaje, no necesitando mayores esfuerzos para mantener la postura y el balance. Por el contrario, Tarrant y col. (1988) utilizando tres densidades distintas (200, 300 y 600 kg/m2) encontraron que al aumentar ésta, aumentaban los valores de CK. La densidad de animales por m2 utilizada en este experimento es cercana a la máxima densidad (600 kg/m2) utilizada por Tarrant y col. (1988).
Respecto a los valores significativamente menores (P<0,05) observados en los animales con descanso en relación a los sin descanso, a la llegada a la PFC (cuadro 1), estaría reflejando el efecto benéfico del período de descanso de 8 h, la ingestión de alimento y de agua y la gran variación individual en la concentración sanguínea de esta variable. La disminución no significativa (P>0,05) en la actividad plasmática de CK del grupo sin descanso en el período de llegada a la PFC y el momento de la sangría se debería al efecto del descanso y a la ingestión de agua en el matadero (gráfico 5). La gran variabilidad en la respuesta individual reflejada en las amplias desviaciones estándar impide obtener conclusiones definitivas acerca de las diferencias entre ambos grupos en este período.
Conclusiones. Existió una gran variación individual en la respuesta de los animales frente al transporte por 36 h. El tratamiento sin descanso provocó un mayor estrés, una mayor tendencia a la movilización grasa y al daño muscular reflejado por las concentraciones de cortisol, ß-HBA y CK al momento de la sangría. Los altos valores de glucosa al momento de la sangría podrían indicar una deficiente técnica de noqueo.
Transportes prolongados, con o sin descanso, como los utilizados en este estudio, conspiran contra el bienestar animal, tal como lo indican las alteraciones observadas en las concentraciones sanguíneas de las variables indicadoras de estrés.
RESUMEN
Se realizó un experimento en invierno (juniojulio) con el objetivo de determinar el efecto de un tiempo de transporte terrestre de 36 h, con y sin descanso, sobre las concentraciones sanguíneas de cortisol, glucosa, ß-HBA, valores de hematocrito y actividad plasmática de CK, en bovinos.
Se emplearon 40 novillitos y vaquillas cruzas de Angus y Hereford procedentes del mismo predio, de DL y 2D de edad y de un peso promedio de 400 kg, divididos al azar en 2 grupos de 20 animales cada uno, los cuales fueron transportados durante 36 h, con y sin un período de descanso de 8 h, después de las primeras 24 h, de viaje. Los animales fueron faenados luego de un período mínimo de 12 h, de reposo en la planta faenadora de carnes (PFC).
Las muestras de sangre se obtuvieron por punción yugular en el predio, la estación de descanso, la PFC y al momento de la sangría. La determinación de cortisol se realizó por radioinmunoensayo (RIA); el VGA mediante microhematocrito; la glicemia mediante el procedimiento para la glucosa GOD-GAP, sin deproteinización (GL 2623, RANDOX,), el ß-HBA mediante la técnica enzimática que utiliza la enzima ß-HBA deshidrogenasa para medir la reducción del NAD+ a NADH; la actividad plasmática de CK mediante el método UV-cinético, a 340 nm y a 37C. Las concentraciones de glucosa, ß-HBA y CK se determinaron con un espectrofotómetro HITACHI 4020. Para el análisis de los resultados se utilizó estadística descriptiva, determinándose la significancia de la diferencia entre las medias mediante el análisis de varianza (ANOVA) o el análisis de Kruskal-Wallis cuando las varianzas no eran homogéneas.
Se observó una gran variación individual en la respuesta al estrés por transporte, reflejado por las grandes desviaciones estándar. De los resultados se desprende que el transporte de bovinos en pie por 36 h, con o sin descanso, es perjudicial para el bienestar de los animales. Esto se reflejó en el aumento significativo de las concentraciones plasmáticas de cortisol, glucosa y CK entre el predio y la PFC y el momento de la sangría. El descanso tuvo un efecto beneficioso sobre la actividad plasmáticas de CK y los valores de VGA y en una menor tendencia a la movilización grasa reflejada por las concentraciones plasmáticas de ß-HBA al momento de la sangría.
_________________________________________
Aceptado: 21.09.2000.
BIBLIOGRAFIA
BUSTOS, V. 1997. Efecto de la administración de acetato de 9-fluoroprednisolona en el destare, rendimiento y pH de la canal de bovinos sometidos a transporte prolongado. Tesis, M.V. Universidad. Austral de Chile, Facultad de Ciencias Veterinarias, Valdivia, Chile.
CABALLERO, S.C:, H.S SUMANO. 1993. Caracterización del estrés en bovinos. Arch. Med. Vet. 1: 15-30.
CHILE. 1992. Ley 19.162. Establece sistema obligatorio de clasificación de ganado, tipificación y nomenclatura de sus carnes y regula funcionamiento de mataderos, frigoríficos y establecimientos de la industria de la carne. Publicada en Diario Oficial 7 de septiembre de 1992.
CHILE, MINISTERIO DE AGRICULTURA. 1993. Reglamento General de Transporte de Ganado y Carne Bovina. Decreto N 240. Publicado en Diario Oficial 26 de Octubre de 1993.
CHILE. Instituto Nacional de Estadísticas. 1997a. VI Censo Nacional Agropecuario.
CHILE. Instituto Nacional de Estadísticas. 1997b. Estadísticas Pecuarias
COCKRAM, M.S., J.E. KENT, P.J. GOODDARD, N.K. WARAN, I.M. McGILP, R.E. JACKSON, G.E. MUWANGA, S. PRYTHERCH. 1996. Effect of space allowance during transport on the behavioural and physiological responses of lambs during and after transport. Anim. Sci., 62: 461 477.
COOPER, C, A., C.O. EVANS, S. COOK, N.C. RAWLINGS. 1995. "Cortisol, progesterone and ß-endorphin response to stress in calves. Can. J. Anim. Sci. 95: 197-201.
CROOKSHANK, H.R., M.H. ELISSALDE, R.G. WHITE, D.C. CLANTON, H.E. SMALLEY. 1979. Effect of transportation and handling of calves upon blood serum composition. J. Anim. Sci. 48: 430-435.
DEAN, G. A., J.A. DEAN, A. BURTON, R. DICKER. 1997. Epi Info, versión 6.03. A Word Processing, Database, and Statistics System for Epidemiology. USD. Incorporated, Stone Mountain, Georgia.
FORREST, J.C., E.D. ABERLE, H.B. HEDRICK, M.D. JUDGE, R.A. MERKEL. 1979. Fundamentos de Ciencia de la Carne. Ed. Acribia, Zaragoza, España.
GALLO, C. 1994. Efecto del manejo pre y post faenamiento en la calidad de la carne. Serie Simposios y Compendios de la Sociedad Chilena de Producción Animal 2: 27-47.
GALLO, C., X. CARMINE, J. CORREA, S. ERNST. 1995. Análisis del tiempo de transporte y espera, destare y rendimiento de la canal de bovinos transportados desde Osorno a Santiago. Resumenes de la XX Reunión Anual de la Sociedad Chilena de Producción Animal: 205 206. Coquimbo, Chile.
GALLO, C. 1996. Consideraciones sobre el manejo antemortem en Chile y su relación con la calidad de la carne. Informativo sobre carne y productos cárneos (edición especial) 21: 27-46.
GALLO, C., M. ESPINOZA; J. CID; J. GASIC. 1998. Pérdidas de peso y características de la canal en bovinos transportados por carretera durante 36 horas, con o sin abrevaje. Resúmenes de la XXIII Reunión Anual de la Sociedad Chilena de Producción Animal: 175-176. Chillán, Chile.
GRANDIN, T. 1994. Farm animal welfare during handling, transport and slaughter. JAVMA, 204: 372 - 377.
HERDT, T.H. 1988 Fuel homeostasis in the ruminant. Veterinary Clinics of North America: Food Animal Practice, 4: 213 231.
HORTON, G.M.J., J.A. BALDWIN, S.M. EMANUELE, J.E. WOHLT, L.R. MCDOWELL 1996. Performance and blood chemistry in lambs following fasting and transport. Anim. Sci., 62: 49-66.
KNOWLES, T.G., P.D. WARRIS, S.N. BROWN, J.E. EDWARDS, P.E. WATKINS, A.J. PHILLIPS. 1997. Effects on calves less than one month old of feeding or not feeding them during road transport of up to 24 hours. Vet. Rec., 140: 116 124.
KNOWLES, T.G. 1999. A review of road transport. Vet. Rec., 144: 197 201.
KNOWLES T.G.,P.D. WARRISS, S.N. BROWN, J.E. EDWARDS 1999. Effects on cattle of transportation by road for up to 31 hours. Vet. Rec., 145: 575 582.
LISTER, D., N.G. GREGORY, P.D. WARRISS. 1981. Developments in meat science. Applied Science Publishers. London.
MATIC, M.A. 1997. Contusiones en canales bovinas y su relación con el transporte. Tesis, M.V. Universidad Austral de Chile, Facultad de Ciencias. Veterinarias. Valdivia, Chile.
MITCHELL, G., J. HATTINGH, M. GANHAO. 1988. "Stress in cattle assessed after handling, after transport and after slaughter". Vet. Rec., 123: 201-205.
SHAW, F.D, R.K. TUME. 1992. The assessment of pre-slaughter and slaughter treatments of livestock by measurement of plasma constituents-A review of recent work. Meat Science, 32: 311-329.
TADICH, N., M. ALVARADO, C. GALLO. 1999. Efecto de 3, 6, 12 y 24 horas de transporte terrestre continuo sobre algunas variables indicadoras de estrés en bovinos. Resúmenes de XXIV Reunión Anual de de la Sociedad Chilena de Producción Animal: 173 174. 27 29 de Octubre, Temuco, Chile.
TARRANT, P.V., F.J. KENNY, D. HARRINGTON. 1988. The effect of stocking density during 4 hour transport to slaugther on behaviour, blood constituents and carcass bruising in friesian steers. Meat Science, 24: 209-222.
TUME, R.K., F.D. SHAW. 1992. Beta-endorphin and cortisol concentration in plasma of blood samples collected during exsanguination of cattle. Meat Science, 31: 211-217.
WARNER, R.D., G.A. ELDRIDGE, C.G. HALPIN, J.L. BARNETT, C.G. HALPIN, D.J. CAHILL. 1986. The effects of fasting and cold stress on dark-cutting and bruising in cattle. Proc. Aust. Soc. Anim. Prod., 16: 383-386.
WARRISS, P.D., S.C. KESTING, S.N. BROWN, L.J. WILKINS. 1984. Recovery from mixing stress in young bulls. Meat Science, 10: 53-68.
WARRISS, P.D. 1990. The handling of cattle pre-slaughter and its effects on carcass and meat quality. Applied Animal Behaviour Science, 28: 171-186.
WARRISS, P. 1992. Animal welfare. Handling animal before slaughter and the consequences for welfare and product quality. Meat Focus International: 135-138.
WARRISS, P.D., S.N. BROWN, T.G. KNOWLES, S.C. KESTIN, J.E. EDWARDS, S.K. DOLAN, A.J. PHILLIPS. 1995. "Effects on cattle of transport by road for up to 15 hours". Vet. Rec., 136: 319-323.
WITTWER, F., H. BÖHMWALD. 1983. Manual de Patología Clínica Veterinaria. Facultad de Ciencias Veterinarias, Universidad Austral de Chile, Valdivia, Chile.