Síndrome Estrés Porcino
Porcine Stress Syndrome
A. M. BONELLI,1 MV.; C. SCHIFFERLI R, 1 MV., MSc, PhD.
1 Depto. de Patología Animal, Facultad de Agronomía y Veterinaria, Universidad Nacional de Río Cuarto, Ruta Nac. 36, Km. 601, CP: 5800, Río Cuarto, Córdoba, Argentina. Tel. :(54)-358-4676200, Correo electrónico: abonelli@ayv.unrc.edu.ar
SUMMARY
The Porcine Stress Syndrome or Malignant Hyperthermia is an inherited autosomal recessive disease, where the susceptible allele is denominated T and the normal stress- resistant allele C. The stress susceptible pigs present death prior to slauhgter and pale, soft and exudative meat in the carcass. The Ryr1 gene mutant in 1843 nucleotide is the cause of Porcine Stress Syndrome. This gene has been studied and discussed widely. At the beginning of the Seventies this gene was important for genetic companies when speculated with the existence of a monogenic variation in the susceptibility to stress, and Eikelenboom and Minkema demonstrated that the Stress Syndrome could be caused by the halothane gas. Most of the studies in positive stress pigs and negative stress pigs showed differences in pH, colour, tenderness and water-holding capacity in the meat. This variation was related to the incidence of pale, soft, exudative meat in the two genotypes. In 1990 MacLennan and collaborators proposed that the ryanodine receptor gene was the gene candidate that predisposed to Malignant Hyperthermia. Fujii in 1991 discovered the mutation responsible for the Porcine Stress Syndrome in the Ryr1 gene, which codifies the calcium release channel of the sarcoplasmic reticulum by the skeletal muscle. The analyses based on DNA (PCR, RFLP) allow to diagnose the three genotypes of the SSP, directing the present genetic selection to obtain resistant pigs to Porcine Stress Syndrome.
Palabras claves: hipertermia maligna, carne PSE, gen ryr1, mutación nucleótido 1843.
Key words: malignant hiperthermia, PSE meat, ryr1 gene, 1843 nucleotide mutation.
INTRODUCCION
En la actualidad, la optimización en la producción porcina y la selección de carne de cerdo de calidad, deben ir acompañadas de la aplicación de la biotecnología del ácido desoxirribonucleico (ADN). La biotecnología del ADN presenta grandes ventajas e innumerables aplicaciones, como son: el chequeo de parentesco, trazabilidad, tipificación de genes de interés en producción o marcadores asociados a caracteres cuantitativos de producción, y detección de enfermedades hereditarias (Haley y col., 1991; Calvo y col., 1997; De Vries y col., 2000). El Síndrome Estrés Porcino (PSS) o Hipertermia Maligna (MH) es una enfermedad hereditaria monogénica recesiva que se caracteriza por un desorden neuromuscular (Shen y col, 1992), con un locus autosomal único, denominado inicialmente gen halotano (HAL) y actualmente llamado gen receptor de la ryanodina "Ryr1" (Dekkers, 1999). El PSS se caracteriza por producir la muerte en los cerdos homocigotos recesivos, así como la aparición de canales con carne pálida, blanda y exudativa (PSE) o carne oscura, dura y seca (DFD), con menor frecuencia, en cerdos homocigotos recesivos y heterocigotos. Esto representa graves pérdidas económicas para la industria porcina, ya que estas carnes no tienen mercado y son decomisadas. Antiguamente, el diagnóstico de esta enfermedad se realizaba con la prueba del halotano. Los avances en el campo de la genética molecular (De Vries y col., 2000) han puesto claramente en evidencia el descubrimiento de la equivalencia entre el PSS y la mutación del gen Ryr1 en el nucleótido 1843 (Fujii y col., 1991). Por medio de la Reacción en Cadena de la Polimerasa (PCR), hoy se diagnostican a los cerdos como sanos (homocigotos dominantes), portadores (heterocigotos) y enfermos (homocigotos recesivos) (Brascamp y col., 1995; Calvo y col., 1997). La estrategia de selección de las empresas genéticas parece ser la eliminación del gen Ryr1 con la mutación 1843 en las líneas materna y paterna (Webb, 1996).
SINONIMIA
Hipertermia Maligna, Muerte Durante el Transporte, Carne de Cerdo, Pálida, Blanda y Exudativa y Necrosis Muscular del Lomo.
ETIOLOGIA
En los últimos 35 años, el sector
productor de cerdos, a nivel mundial, se ha esforzado en seleccionar cerdos
magros. Sin embargo, a partir de la década de los sesenta, se observó
que los resultados de selección iban unidos a una alta mortalidad por
estrés (Calvo y col. 1997). Se detectó que
la selección de reproductores con mejores características magras
y mayor desarrollo muscular implicaba animales enfermos o portadores de la
enfermedad PSS, trasmitiendo a la descendencia el carácter. Hoy se
conoce que el PSS presenta un modelo de herencia mendeliana recesiva simple.
La enfermedad está controlada por un gen recesivo llamado gen receptor
de la ryanodina, Ryr1 (Fujii y col., 1991), anteriormente
denominado gen Halotano (Dekkers, 1999) con un locus autosomal
único que se encuentra ubicado en el cromosoma 6 del cerdo. El gen
Ryr1 está constituido por 15.105 nucleótidos y tiene dos expresiones
o alelos: el alelo normal es dominante sobre el alelo mutado. El control genético
de la enfermedad implica que la expresión del alelo recesivo, en el
individuo, ha de tener la condición homocigótica recesiva, de
forma que en condición heterocigótica, el individuo es normal,
permaneciendo el alelo mutado escondido, pudiendo trasmitirse a las siguientes
generaciones. De esta forma los individuos pueden presentar tres genotipos
diferentes a saber: 1. Individuos normales: sanos no portadores, homocigotos
dominantes (CC).
2. Individuos portadores: sanos, portadores del alelo mutado T a la descendencia,
transmisores del alelo mutado, heterocigotos (CT).
3. Individuos enfermos: trasmisores del alelo mutado, homocigotos recesivos
(TT), (Calvo y col., 1997). Ambos alelos exhiben penetrancia
incompleta (Minkena y col., 1977). El canal liberador de
calcio (CRC) es codificado por tres genes denominados Ryr1, Ryr2 y Ryr3, con
expresión de tejido específico. Solamente el gen Ryr1 se expresa
en el músculo esquelético y también en el cerebro, particularmente
en las células cerebelares de Purkinje. El gen Ryr2 se expresa en el
corazón, endotelio y mayormente en el cerebro. El gen Ryr3 se manifiesta
en el músculo liso, epitelios, y cerebro (O' Brien,
1995).
ORIGEN
EPIDEMIOLOGIA
El PSS ocurre en todo el mundo, pero existe una considerable variación en cuanto a la frecuencia en las razas y regiones. En algunos países de Europa la prevalencia de este síndrome ha aumentado durante los últimos años. Hoy constituye un problema importante en la producción porcina. Tal hecho ha dependido de la selección negligente o descuidada respecto a este síndrome en los programas de mejoramiento genético, en especial los programas de selección basados puramente en características de rendimiento y producción. La enfermedad probablemente ocurra en todas las razas porcinas, pero es más frecuente en las seleccionadas por una fuerte musculatura, con escasa grasa dorsal y más carne. Es sobre todo frecuente en las razas Pietrain, Poland China y Landrace, para las cuales, se ha incluido en el índice de selección, un sistema de puntos en cuanto a musculatura, ritmo de crecimiento, conversión de alimento, y espesor de grasa dorsal (Blood y col., 1988).
Los factores estresantes que pueden desencadenar el PSS son: 1. Estresantes físicos: transporte a temperatura y/o humedad ambiental alta, ejercicio físico durante la carga y descarga, privación de comida y agua, alojamiento y transporte prolongado (Shen y col., 1992; Manteca, 1998). 2. Estresantes sicológicos: mezcla de animales, ambiente nuevo, manejo brusco previo, durante y posterior al transporte de los animales y apareamiento. (Shen y col., 1992; Manteca, 1998). 3. Estresantes farmacológicos: anestésicos como halotano, metoxiflurano, cloroformo, enflurano, y fluroxeno. Los relajantes musculares despolarizantes como la succinilcolina y los agonistas a - adrenérgicos también pueden iniciar o potenciar el síndrome (Shen y col., 1992; O'Brien, 1995).
Está ampliamente aceptado que el cerdo es una de las especies más sensibles al estrés del transporte. Esto es consecuencia de al menos tres factores a saber: 1. Termorregulación: si bien el cerdo tiene glándulas sudoríparas funcionales, su capacidad de perder calor mediante sudor es prácticamente nula. Esto hace que sea particularmente sensible a las temperaturas elevadas, especialmente cuando otros mecanismos de pérdida de calor, tales como la vasodilatación periférica, se ven comprometidas a consecuencia de la respuesta a estrés. 2. Estrés social: el cerdo es un animal social, cuyos grupos se organizan en torno a una jerarquía. Esto se produce mediante interacciones agresivas. Al mezclar cerdos procedentes de grupos diferentes, la jerarquía debe establecerse otra vez. Las agresiones pueden causar lesiones en los animales y son por otra parte, un factor estresante intenso. 3. Sensibilidad al estrés: los animales genéticamente sensibles a estrés padecen una alteración en la membrana del retículo sarcoplásmico de la fibra muscular estriada. A consecuencia de ello, la concentración citoplasmática de calcio, responsable del mecanismo de contracción muscular, permanece anormalmente elevada durante un período de tiempo excesivamente largo. Esto hace que los animales sensibles a estrés, tengan dificultades a la hora de controlar la contracción muscular. En situaciones estresantes los animales homocigotos recesivos pueden sufrir un cuadro de hipertermia y acidosis frecuentemente fatal; tanto los homocigotos recesivos como los heterocigotos presentan a menudo carne pálida, blanda y exudativa, dependiendo de las reservas de glucógeno muscular (Webb, 1996; Manteca, 1998).
La prevalencia del PSS en la población porcina puede determinarse mediante las pruebas de halotano, creatinina quinasa, y ADN. A comienzos de la década del 90 se identificó la mutación del PSS por medio de la PCR, presentando la siguiente distribución racial: 97% en Pietrain, 80% en Poland China, 37% en Landrace, 22% en Large White, Duroc y Hampshire; y 17% en Yorkshire. La mutación también se identificó en las razas Berkshire y Chester White. El PSS tenía mayor prevalencia en Inglaterra, que en Norteamérica y más prevalencia en Estados Unidos que en Canadá (O´Brien, 1995).
METABOLISMO DEL MUSCULO ESQUELETICO
El músculo esquelético cumple varias funciones en el organismo. La función primaria del tejido muscular es producir movimiento y postura. Una notable característica del músculo esquelético es su capacidad para cambiar con rapidez de un estado relajado a uno activo. Los procesos cronológicos que se dan durante la actividad del músculo esquelético incluyen: transmisión colinérgica de una señal de despolarización proveniente de una neurona motora espinal, propagación de este potencial de acción en la superficie de la membrana plasmática hacia la célula muscular por medio de los túbulos T, liberación de calcio por medio del canal liberador de calcio a partir de la reserva intracelular en el retículo sarcoplásmico hacia el citoplasma de la célula muscular, unión del calcio con los filamentos de la troponina, inicio de la contracción muscular, activación de vías glicolíticas, síntesis de ATP, contracción muscular, ingreso de calcio al retículo sarcoplásmico por medio de la bomba de calcio ATPasa y relajación muscular (Ruckebush y col; Calvo y col., 1997). El músculo esquelético es el tejido cuantitativamente más importante del organismo, representando el 60 % del peso de la canal para un cerdo de 105 kg de peso vivo; está constituido mayoritariamente por agua (73%), proteínas (20%), lípidos (1-6%) y glúcidos (1-2%). Asimismo, las fibras musculares constituyen el 75 a 90 % del volumen muscular y el resto está integrado por tejido adiposo, conjuntivo, vascular y nervioso. Las fibras musculares en el cerdo se clasifican en: tipo I (lentas, oxidativas y rojas); tipo II A (rápidas, intermedias y rojas) y tipo II B (rápidas, glicolíticas y blancas). Los músculos del cerdo contienen los tres tipos de fibras, pero en proporciones variables, según la unidad motora. Así, en el músculo Longissimus dorsii (LD) del cerdo, el contenido relativo de las fibras I: IIA: IIB (8: 8: 84), en comparación al de la vaca (50: 40: 10), predispone a la carne de cerdo a una mayor incidencia de PSE y DFD (Barton Gade, 1997). Asimismo, la tendencia hacia animales con crecimiento rápido, con menos grasa y masas musculares desarrolladas favorecen la mayor incidencia de las fibras blancas en perjuicio de las rojas. De esta forma, en los músculos LD, biceps femoral, semimembranoso, semitendinoso, y glúteo se observa una disminución del color rojo, acompañado frecuentemente por un nivel más alto de humedad superficial, aunque sin llegar a fenómenos de verdadera exudación.
FISIOPATOLOGIA
En el cromosoma 6 del cerdo se encuentra el gen Ryr1, que codifica una proteína denominada receptor de la ryanodina o canal liberador de calcio del retículo sarcoplásmico del músculo esquelético estriado (CRC), con un peso molecular de 564.743 daltons (O'Brien, 1995). Fujii y col. (1991) detectaron el punto de mutación en el gen Ryr1 que se asoció con MH en 6 razas de cerdos domésticos (Bauerová y col., 1995). Esta mutación consiste en la sustitución de una timina por una citosina en la posición 1843 de la secuencia de ADN complementario del gen Ryr1, que lleva a la sustitución de una arginina por cisteína en la posición 615 del CRC del animal sensible. La presencia en el individuo de la condición homocigótica en el gen Ryr1 (TT) provoca una alteración en la liberación de calcio en el retículo sarcoplásmico del músculo esquelético estriado. El retículo sarcoplásmico es el principal regulador de la concentración de calcio en el músculo esquelético. A través del CRC, el calcio sale al citoplasma de la célula muscular uniéndose a los finos filamentos de troponina e iniciando así la contracción del músculo. El calcio se une a una fosforil quinasa que activa las vías glucolíticas (glucógeno - glucosa) y de síntesis de ATP necesarias para la contracción muscular. Por medio de la bomba de calcio ATPasa, el calcio ingresa en el retículo sarcoplásmico iniciándose la relajación. Ambos mecanismos mantienen el balance energético del miocito. En un CRC anormal se produce la liberación de calcio en forma continua, existiendo un aumento de la concentración en el calcio citoplasmático del miocito. El metabolismo aeróbico, la glucogenólisis y la glucólisis se incrementan, agotando el ATP, la glucosa y el oxígeno, produciéndose un exceso de dióxido de carbono, ácido láctico, potasio y calor en la sangre, además de un desorden en el balance de iones intra y extracelular, llevando asimismo a un acúmulo de agua en la célula. La asociación marcada entre respuesta a estrés fisiológico, con hipercatecolemia, produce paro cardíaco. El estado de contracción permanente de la célula muscular provoca hipertrofia muscular, con el consiguiente aumento del desarrollo muscular de la canal (Reiner, 1993; ; O'Brien, 1995; Calvo y col., 1997).
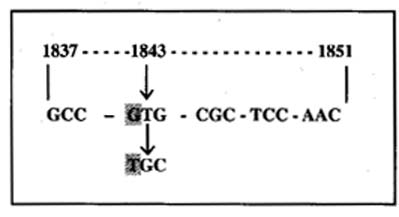
Figura 1. Secuencia de nucleótidos
del gen Ryr1 que muestra la mutación en la posición 1843 (Fujii
y col., 1991).
Sequence of nucleotides within the Ryr1 gene showing mutation
at position 1843 (Fujii y col., 1991).
MANIFESTACIONES CLINICAS
Hipertermia maligna (HM). Puede ser inducida en animales susceptibles por anestésicos volátiles y potentes como el halotano o por administración de succinilcolina. Se caracteriza por incremento del metabolismo del músculo durante la anestesia con rigidez muscular, acidosis láctica y aumento manifiesto del índice metabólico basal, incremento del consumo de oxígeno, producción de dióxido de carbono, hipertermia grave, taquicardia, taquiarritmia y muerte. Una vez desarrollado el síndrome es irreversible, representando un evidente riesgo la anestesia en los cerdos, la cual, puede prevenirse por medicación previa con dandroleno. Se ha usado también como método para determinar la susceptibilidad al estrés en programas de selección genética (Blood y col., 1988; Fujii y col., 1991; O'Brien, 1995).
Carne pálida, blanda y exudativa (PSE). En los cerdos susceptibles a estrés, con frecuencia, la carne es de calidad inferior después del sacrificio, ya que se observa a la inspección, pálida, blanda y exudativa. Esto se relaciona con glucogenólisis y glucólisis excesiva después de la muerte, producción de ácido láctico y caída rápida del pH del músculo, alcanzando valores inferiores a pH 6, antes de la primera hora después del sacrificio, con despigmentación y merma en el agua de fijación. En el músculo afectado, rápidamente se produce rigidez cadavérica luego del sacrificio, pero a continuación disminuye, de modo que en la carne de la canal, se observa excesiva exudación. La carne afectada tiene pH menor a 6 y generalmente temperatura de 41°C o mayor, 45 minutos después del sacrificio, en oposición al pH normal superior a 6 y temperatura menor de 40 °C. Asimismo, la carne tiene sabor desagradable y cualidades netamente inferiores respecto a la cocción y elaboración, no aceptando el encurtido con facilidad (Blood y col., 1988; Webb, 1996; Calvo y col., 1995). En Estados Unidos se ha estimado que se pierden unos $ 100 millones anuales debido a carne PSE (Kauffman y col., 1998). La existencia de carne DFD aparece menos frecuentemente en animales muertos por PSS. Esto se produce por un pH superior a 6, después de haber transcurrido 24 horas del sacrificio, reteniendo mayor cantidad de agua y deteriorándose la canal con facilidad (; Webb, 1996; Calvo y col., 1997). Los cerdos resistentes al estrés, homocigotos dominantes, típicamente no producen carne PSE, pero sí lo manifiestan comúnmente los homocigotos recesivos (Monin y col., 1999).
Necrosis del músculo del lomo, músculo dorsal o enfermedad de la banana. Cuando los cerdos superan un estrés grave durante más de 2 horas, se produce el cuadro clínico de la llamada necrosis aguda del músculo dorsal (Otto Eich, 1991). El síndrome agudo dura 2 semanas aproximadamente y se caracteriza por inflamación y dolor sobre los músculos del lomo. Se inflama un músculo dorsal largo, pero raras veces ambos. El dorso del animal enfermo se curva hacia arriba, con convexidad hacia el lado afectado. Hay renuncia de movimiento por parte del animal. La inflamación y el dolor mejoran después, pero hay atrofia del músculo afectado y rigidez prominente de la columna vertebral, si bien puede ocurrir alguna regeneración al cabo de varios meses. En los casos agudos, es muy frecuente la muerte. Suele observarse este síndrome en adultos jóvenes con peso de 75 a 100 kg. La forma leve puede pasar inadvertida excepto en pacientes que adoptan decúbito cerca del lugar en que comen. En la forma grave, los cerdos afectados, pueden asumir la posición de perro sentado con el lomo arqueado (Blood y col., 1988; Otto Eich, 1991). Los efectos de la genética y la sensibilidad al halotano sobre la calidad de la canal y de la carne, se han revisado en distintos estudios (De Vries y col., 1999). Los cerdos halotano positivos en relación a los cerdos halotano negativos, incluyendo el homocigoto dominante, y heterocigoto, se diferencian por tener:
- Menor crecimiento y apetito.
- Canales más cortas, mejor conformadas, y con mayor rendimiento.
- Menor espesor de grasa dorsal y mayor porcentaje de magro.
- Mayor área y profundidad del músculo LD.
- Elevada incidencia de carne PSE.
- Reducción del tamaño de camada.
Wysocki y col. (1998) estudiaron en 35 verracos de un centro de inseminación polaco la influencia del gen Ryrl sobre la calidad del semen y su capacidad de conservación en estado líquido arribando a las siguientes conclusiones:
1. El volumen y número de espermatozoides totales en el eyaculado son significativamente menores en los verracos homocigotos recesivos. 2. El rápido deterioro de la integridad de la membrana y la disminución de la motilidad desaconseja que el esperma de los verracos homocigotos recesivos sea utilizado para conservarlo a largo plazo en medio líquido. 3. El semen conservado de los verracos homocigotos recesivos tiene baja fertilidad. Desde un punto de vista económico, el semen de dichos verracos no debe ser utilizado para la inseminación artificial.
Warnants y col. (1993) advirtieron que los cerdos homocigotos recesivos que tienen una buena musculatura, se ven frecuentemente acompañados por una inferior calidad de la carne, menor fertilidad, y mayores niveles de mortalidad en lactancia.
DIAGNOSTICO
Existen varios métodos para predecir la susceptibilidad de los cerdos al PSS.
Prueba de halotano. El primer diagnóstico del PSS se realizó a partir de la prueba del halotano, debido a la relación existente entre el anestésico halotano y el PSS (Eikelenboom y Minkema, 1974). Esta prueba es sumamente confiable para la identificación de los cerdos homocigotos recesivos. Se conoce que el diagnóstico por halotano detecta más del 90% de los homocigotos y menos del 10 % de los heterocigotos para el defecto PSS (Shen y col., 1992). Este método tiene el inconveniente que no diferencia entre animales sanos no portadores y sanos portadores e identifica a algunos animales enfermos homocigotos recesivos como normales (Calvo y col., 1997). Los cerdos susceptibles a estrés son sensibles al halotano a las 8 semanas de edad. Si el peligroso anestésico se retira inmediatamente después de la aparición de signos evidentes de rigidez en las extremidades y antes de la aparición de la hipertermia fulminante, la mortalidad del método es mínima. El método consiste en administrar por vía inhalatoria el anestésico halotano y observar la reacción del animal. Los cerdos que permanecen no reactivos durante un período de 5 minutos se consideran normales. Los cerdos se adormecen sin aparecer contracturas ni espasmos musculares. Los animales enfermos manifiestan contracturas y espasmos musculares (Blood y col., 1988; Grobet y col., 1992; Calvo y col., 1997).
Niveles de creatina - cinasa en la sangre. Estos niveles son más altos en cerdos susceptibles a estrés y se consideran útiles para la identificación de los animales con tal susceptibilidad. Puede utilizarse la prueba de la creatina - cinasa como criterio selectivo para estimar la resistencia a estrés y la calidad de la carne. Para ejecutarla se somete primero a los cerdos a una prueba de ejercicio normal y se toman muestras de sangre 8 a 24 horas después con el objeto de determinar los niveles de creatina - cinasa. Es buena la correlación entre dichos niveles y la prueba del halotano. Se observa también un incremento en los niveles de enzimas séricas en porcinos durante el transporte de la granja al matadero. Sin embargo, no se encuentran niveles elevados de creatina - cinasa en el suero de todos los cerdos con el cuadro de PSE. La prueba inicial se ha modificado de modo que la sangre puede colectarse en forma de gotas sobre un papel filtro y se envía por correo al laboratorio para identificación por una técnica bioenzimática. El método de la luciferasa para determinación de creatina - cinasa en sangre entera se considera un procedimiento discriminativo útil para el PSS (Blood y col., 1988).
Tipificación de sangre. Este método se usa para la identificación de cerdos susceptibles. En el cromosoma 6 del cerdo se ha identificado una región con 4 loci, los cuales contienen los genes responsables de las variantes de las enzimas 6 fosfogluconato deshidrogenasa y fosfo isomerasa. El sistema del grupo sanguíneo H es determinado por uno de los loci y la sensibilidad al halotano lo es por genes de un locus de esta región, la cual tiene un interés muy particular, debido a que se ha comprobado, una conexión muy estrecha entre ella, con características muy importantes en la carne, como lo es el PSS. De este modo, puede utilizarse el grupo sanguíneo para descubrir cerdos sensibles y portadores a PSS (Blood y col., 1988).
Carne pálida blanda y exudativa. Esta característica se valora mediante un índice de calidad de carne que combina color, pH a las 24 horas después de muerto y capacidad de retención de agua. Casi todas estas pruebas predicen fácilmente los ejemplos más graves del PSS, pero no son lo bastante precisas para identificar tendencias hacia el empeoramiento, lo cual restringe su valor en los programas de reproducción.
Prueba de contracción por cafeína y halotano. Se realiza a partir de biopsias musculares de animales sospechosos. Esta técnica es muy usada en humanos para diagnosticar MH. Las muestras de músculo son colocadas en solución salina normal oxigenada y conectadas a un tensiómetro que mide la fuerza de desplazamiento. La tensión isométrica es grabada por un polígrafo durante la exposición a 5% de halotano incrementándose progresivamente la cantidad de cafeína. La concentración de cafeína en 1 gramo, incrementa la tensión. Cuando la concentración de cafeína es menor que 4 mM y el músculo desarrolla más de 0.5 de tensión se expone al halotano considerándose a los cerdos MH susceptibles. Los heterocigotos tienen resultados intermedios (Shen y col., 1992; O' Brien, 1995).
El uso de técnicas del ADN recombinante, como la PCR y los polimorfismos de la longitud de los fragmentos de restricción (RFLP) (Fujii y col., 1991) y polimorfismos de conformación de cadena simple (SSCP) (Bauerová y col., 1995), permiten la detección de animales sanos, enfermos y portadores con una seguridad del 100 %. Los análisis de ADN se realizan directamente sobre el individuo del que se desea estudiar, sin posibilidad de error.
PCR-RFLP: básicamente el método de diagnóstico desarrollado consiste en la amplificación específica de un fragmento del gen Ryr1, que contiene la mutación, mediante la técnica de PCR. Posteriormente se realiza una digestión con enzimas de restricción y se visualizan los resultados mediante electroforesis en gel de agarosa, diferenciándose claramente los tres genotipos (Calvo y col., 1997).
PCR-SSCP: a diferencia de PCR-RFLP es un método que no requiere enzima de restricción y gel de agarosa. El costo del diagnóstico se reduce a la mitad, es más fácil y rápido que el método PCR-RFLP. Los fragmentos amplificados son visualizados en una corrida electroforética en un gel de acrilamida a 3 °C (Nakajima y col., 1996).
HALLAZGOS A LA NECROPSIA
En el PSS se observa siempre rigidez cadavérica, posterior a la muerte, con putrefacción de carne en la canal más rápidamente que en condiciones normales. Durante la necropsia se comprueba que las vísceras están congestionadas y hay casi siempre aumento del líquido pericárdico con presencia de congestión pulmonar y edema. Los músculos, especialmente glúteo medio, bíceps crural y dorsal largo están pálidos y blandos, lo cual puede ser manifiesto inmediatamente después de la muerte, pero alcanza un máximo al cabo de unas 2 horas. Por examen histopatológico se comprueba degeneración hialina del músculo esquelético y cardíaco. Se aprecian cambios floculares y hialinos. En los casos crónicos hay fagocitosis, mineralización, y regeneración de células musculares con fibrosis (Blood y col., 1988).
TRATAMIENTONo suele emprenderse tratamiento en los síndromes agudos, pero sí se dispone de varios medicamentos para la protección de los cerdos contra la MH inducida por fármacos. En efecto, una combinación de acepromacina y droperidol retardará el comienzo o evitará la aparición de MH inducida por halotano y dandrolene. La necrosis aguda del lomo se trata satisfactoriamente con isopirín y fenilbutazona (Blood y col., 1988).
CONTROL
El control de la enfermedad se basa en la reducción del estrés impuesto sobre los cerdos, manejo apropiado de los animales y selección en contra del alelo mutado.Recomendaciones propuestas para disminuir la incidencia de muertes por estrés en el transporte.
1. Densidad de animales durante el transporte: aunque es difícil dar recomendaciones precisas sobre la densidad máxima de animales durante el transporte, los trabajos coinciden en que menos de 0.35m2 por animal es desaconsejable. Algunos autores, recomiendan densidades considerablemente superiores (0.5 - 0.55 m2 por animal). El diseño del camión es igualmente importante y se deben utilizar sistemas de compartimentación que permitan transportar a los animales en grupos de 6 a 8 animales (Manteca, 1998). El Instituto Americano de la Carne (American Meat Institute, 1991) recomendó para cerdos de 90 kg una densidad de 0.32 m2 en invierno y 0.37 m2 en verano; y para cerdos de 113 kilos una densidad de 0.40 m2 en invierno a 0.46 m2 en verano.
2. Temperatura y ventilación: evitar el transporte durante las horas más calurosas del día en verano y asegurar una ventilación adecuada durante el transporte para reducir las muertes y evitar la disminución en la calidad de la carne. Igualmente, la instalación de duchas en los camiones tiene efectos marcadamente positivos (Manteca, 1998). Cuando la temperatura supera los 15 °C se recomienda usar arena o aserrín mojado para mantener frescos a los cerdos. Si pasa de los 23 °C hay que rociarlos con agua antes de embarcarlos en granjas y ferias. En épocas de calor no cubrir el piso con paja. Cuando haya menos de 15 °C se podrá usar paja o aserrín seco (American Meat Institute, 1991).
3. Rampa de carga y descarga de los animales: cuando la carga y descarga de animales se realiza por rampas, estas no deberían tener una inclinación superior al 15 %. Es importante señalar, por otra parte, la conveniencia de diseñar granjas de cerdos, de modo que el desplazamiento de los animales hasta el camión pueda realizarse con la mayor facilidad posible. Finalmente, debe evitarse, en lo posible, el tratamiento brusco a los animales, durante la carga y descarga (Manteca, 1998).
4. Alimentación y agua antes y durante el transporte: 12 horas de ayuno son suficientes, aunque trabajos recientes sugieren la posibilidad de que el consumo de una cantidad reducida de pienso, poco antes del transporte, podría tener efectos beneficiosos. Por otra parte, los animales deberían tener acceso al agua hasta inmediatamente antes del transporte (Manteca, 1998). Finalmente, de acuerdo con la legislación de la Comunidad Europea, no deben transcurrir, nunca más de 24 horas sin que los animales sean alimentados y abrevados.
5. Sistema de conducción. Una conducción cuidadosa, que evite cambios súbitos de velocidad y virajes bruscos tienen una considerable influencia positiva desde el punto de vista de reducir las pérdidas económicas como a la hora de aumentar el bienestar de los animales (Manteca, 1998).
6. Puede lograrse el control final del PSS mediante selección genética, seleccionando en contra del alelo mutado, es decir, eliminando animales homocigotos recesivos y heterocigotos. De esta forma la prevalencia de PSS se disminuiría en un 65%, manteniendo las condiciones de manejo anterior a la faena. Se desarrolló una línea de machos Pietrain, libre del gen del estrés, de muy buena calidad de carne, comparado con animales de la misma raza homocigotos recesivos, con doble copia del gen mutante. Esta nueva línea, presenta rendimientos de carne magra y conformación equivalentes a las líneas halotano positivas (Gispert y col., 2000). Es lógico, pues, considerar el genotipo de los animales cuando se negocia la integración de los eslabones de la cadena productiva. En la actualidad, las empresas de mejora genética prueban la calidad de la canal en sus cruzamientos comerciales (Gispert y col., 2000).
7. Otra norma de selección más conservadora que la anterior plantea la cruza de cerdas madres resistentes a estrés con verracos portadores, para así mantener la musculatura y el porcentaje de magro en las reses porcinas (Calvo y col., 1997).
RESUMEN
El Síndrome Estrés Porcino o Hipertermia Maligna es una enfermedad hereditaria autosómica recesiva, de penetrancia incompleta donde el alelo mutado es denominado T y el alelo resistente al estrés C. Los cerdos susceptibles al estrés presentan muerte súbita previa al sacrificio o carne pálida, blanda y exudativa en la canal. El gen de la ryanodina mutado en el nucleótido 1843 causa el Síndrome Estrés Porcino. Este gen se ha estudiado y discutido ampliamente. A principios de los años 70 este gen se tornó importante para las empresas de genética, cuando se especuló con la existencia de una variación monogénica en la susceptibilidad al estrés. Eikelenboom y Minkema (1974) demostraron que el Síndrome de Estrés se podía provocar con el gas halotano. La mayoría de los estudios entre cerdos estrés positivo y cerdos estrés negativo mostraron diferencias en pH, color, terneza y capacidad de retención de agua en la carne. Esta variación estaba directamente relacionada con la incidencia de carne pálida, blanda y exudativa en los dos genotipos. Mac Lennan y col. (1990) propusieron que el gen de la ryanodina era el gen candidato que predisponía a la Hipertermia Maligna. Fujii y col. (1991) descubrieron la mutación responsable del Síndrome Estrés Porcino en el gen de la ryanodina, que codifica el canal liberador de calcio del retículo sarcoplásmico del miocito esquelético. Los análisis basados en ADN (PCR, RFLP) permiten diagnosticar los tres genotipos del SSP, dirigiendo la selección genética actual a conseguir cerdos resistentes al Síndrome Estrés Porcino.
BIBLIOGRAFIA
AMERICAN MEAT INSTITUTE. 1991. Washington, DC. Recomendaciones para el manejo de animales en las plantas de faena.
BARTON GADE, P. A. 1997. In: Manipulating pig production VI. Ed. P.D. Cranwell. Australasian Pig Sci. Assoc. 100-123.
BAUEROVA, M., D. VASICEK, P. UHRIN, P. CHRENEK, J. MLYNEK, J. BULLA. 1995. Detection of malignant hyperthermia in pigs by DNA-test using plucked hair as a source of DNA. Pig News and Information. 16: 109-111.
BLOOD, D. C., J. A. HENDERSON, O. M. RADOSTITS. 1988 Medicina Veterinaria. Interamericana. México, D.F. 1352-1356.
BRASCAMP, E. W., C. S. HALEY, M. A. M. GROENEN, L. L. G. JANSS. 1995. PiGMaP: gene mapping and its contribution to meat production and meat quality parameters. Pig News and Information.16: 41-46.
CALVO, J. H., R. OSTA, E. GARCIA-MURO, P. ZARAGOZA. 1997. Síndrome de estrés porcino: aplicación y ventajas de la PCR para su diagnóstico. Med. Vet.14. 2: 110-113.
DEKKERS, J. C. M. 1999. Optimizing strategies for selection on major genes. Plant & Animal Genome VII held at St. Diego, CA. 1-16.
DE VRIES, A. G., L. FAUCITANO, A. SOSNICKI, G. S. PLASTOW. 1999. New developments in guaranteeing the optimal sensory quality of meal. Ed. F. Toldrá y D.J.Troy. Fundación Vaquero, España. 73-89.
DE VRIES, A. G., A. SOSNICKI, G. S. PLASTOW. 2000. Aplicación de nuevas tecnologías para la selección de carne de cerdo de calidad. ANAPORC. 202: 19-24.
EIKELENBOOM, G., Y. D. MINKEMA. 1974. Prediction of pale, soft, exudative muscle with an non-lethal test for halothane induced porcine malignant hyperthermia syndrome. Neth. J. Vet. Sci. 99: 421-426.
FUJII, J., K. OTSU, F. ZORZATO, S. DE LEON, V. K. KHANNA, J. E. WEILER, P. J. O' BRIEN, D. H. MACLENNAN. 1991. Identification of mutation in porcine ryanodine receptor associated with malignant hiperthermia. Science. 253: 448-451.
GISPERT, M., M. D. GUARDIA, A. DIESTRE. 2000. La calidad de la carne porcina. ANAPORC. 200: 76-80.
GROBET. L., R. HANSET, C. DASNOIS. 1992. Réponse au test a l halothane et génotype au locus Ryr1 du récepteur a la ryanodine chez des porcs croises Pietrain. Ann. Méd. Vét. 136: 249-257.
HALEY, C., J. BROWN, H. MCQUEEN, S. COUPERWHITE, A. ARCHIBALD. 1991. The pig gene mapping project (PiGMaP). Pig News and Information. 14: 13-16.
KAUFFMAN, R. G., R. L. J. M. VAN LAACK, R.L. RUSSELL, E. POSPIECH, C.A. CORNELIUS, C. E. SUCKOW, M.L. GREASER. 1998. Can pale, soft, exudative pork be prevented by postmortem sodium bicarbonate injection ? J. Anim. Sci. 76: 3010-3015.
MACLENNAN, D. H., C. DUFF, F. ZORZATO, J. FUJII, M. PHILLIPS, R. KORNELUK, W. FRODIS, B. BRITT, R. G. WORTON. 1990. Rianodine receptor gene is a candidate for predisposition to malignant hypertermia. Nature. 343: 559-561.
MANTECA., X. 1998. Bienestar animal. XIX Symposium ANAPORC. EXPOAVIGA. 265-272.
,MINKENA D., G. EIKELEMBOOM, P. VAN ELDIK. 1977. Inheritance of MHS- susceptibility in pigs. Proceedings of the Third International Conference on Production Disease in Farm Animals, Wageningen, Netherlands, Pudoc. 203-220.
MONIN, G., C. LARZUL, P. LE ROY, J. CUCIOLI, J. MOUROT, S. ROUSSET-AKRIM, A. TALMANT, C. TOURAILLE, P. SELLIER. 1999. Effects of the halothane genotype and slaughter weight on texture of pork. 77: 408-415.
NAKAJIMA, E., T. MATSUMOTO, R. YAMADA, K. KAWAKAMI, K. TAKEDA, A. OHNISHI, M. KOMATSU. 1996. Use of a PCR-single strand conformation polymorphism (PCR-SSCP) for detection of point mutation in the swine rianodine receptor (Ryr1) gene. J. Anim. Sci. 74: 2904-2906.
O' BRIEN, P. J. 1995. The causative mutation for porcine stress syndrome. Food Animal. 257-269.
OTTO EICH, K. 1991. Enfermedades del cerdo en explotación intensiva. EDIMD. 57-58.
REINER, G. 1993. A new physiological pathway controlling muscle growth and its potencial relevance for pig production. Pig News and Information. 14: 123-125.
RUCKEBUSCH, Y., L. PHANEUF, R, DUNLOP. 1994. Fisiología de pequeñas y grandes especies. Editorial El Manual Moderno de C.V. 527-547.
SHEN, H., R. LAHUCKY, L. KOVAC, P. J. O' BRIEN. 1992. Comparison of Hal gene status with 31P NMR-determined muscle metabolites and with Ca sequestration activity of anoxia - challenged muscle from pigs homozygous and heterozigous for porcine stress syndrome. Pig News and Information. 13: 105-109.
WARNANTS, N., W. EECKHOUT, CH.V.J. BOUCQUE. 1993. Serum pyruvate kinasa and its relation to stress susceptible in pigs. Anim. Breed. Genet. 110: 357-362.
WEBB, A.J. 1996. Future challenges in pig genetics. Pig News and Information. 17: 11-16.
WYSOCKI, P., F. SAIZ CIDONCHA, J. STRZEZEK. 1998. Influencia de la mutación del gen receptor de la ryanodina (Ryr1) en verraco sobre la calidad del semen y su capacidad de conservación en estado líquido. ANAPORC. 182: 144-154.
Aceptado: 07.08.2001