FIGURA 1. Dolly: El primer mamífero clonado por transferencia nuclear desde una célula adulta.
Dolly: The first mammal ever cloned by nuclear transfer from an adult mammary cell.
Arch. Med. Vet., Vol. XXXVI N° 2, 2004, pp. 105-117
REVISIONES BIBLIOGRAFICAS
Animales transgénicos: pasado, presente y futuro
Transgenic animals: past, present and future
R. FELMER, B.Q, PhD.
Unidad de Biotecnología, Instituto de Investigaciones Agropecuarias INIA-CRI Carillanca, Casilla 58-D, Temuco, Chile (rfelmer@carillanca.inia.cl)
Resumen
Desde la demostración del primer animal transgénico en 1980, la ingeniería genética ha revolucionado todos los aspectos de la investigación biológica y biomédica. Desde entonces, se ha logrado la generación de varios tipos de animales transgénicos, incluyendo vacas, cerdos, ovejas, cabras y conejos. Hasta muy recientemente, la generación de animales transgénicos involucraba la microinyección de pequeñas cantidades de ADN en el pronúcleo de un embrión al estado de dos células, técnica conocida como microinyección pronuclear. El descubrimiento de que los animales podrían ser clonados mediante transferencia nuclear de células mantenidas en cultivo abrió las puertas para realizar recombinación homóloga en estas especies. Esto podría tener importantes implicaciones especialmente para los propósitos del descubrimiento de nuevas drogas, para mejorar características productivas de los animales, en la clonación de cerdos como fuentes de órganos para trasplantes y en la producción de proteínas farmacéuticas. En esta revisión se discuten los avances que ha tenido la tecnología para la generación de animales transgénicos y los beneficios de la transferencia nuclear como una nueva ruta para la generación de animales transgénicos de granja.
Palabras clave: animales transgénicos, transferencia nuclear, clonación.
Summary
Since the initial demonstration in 1980 that a transgenic animal could be generated harbouring a transgene from a different species, genetic engineering has revolutionized all aspects of fundamental biological and biomedical research. Since then, much has been accomplished in the generation of various types of transgenic animals, including cows, pigs, sheeps, goats and rabbits. Until recently, genetically modified livestock could only be generated by pronuclear injection. The discovery that animals can be cloned by nuclear transfer from cultured somatic cells means that it is now possible to achieve gene targeting in these species. This may have important implications for the purposes of drug discovery research, to improve animal health and productivity, in cloning pigs as a source donor for xenotransplantion, and in the production of pharmaceutical proteins. This review discusses the development of transgenic technology and the potential benefits emerging from nuclear transfer as a novel route for the generation of transgenic livestock.
Key words: transgenics, nuclear transfer, cloning.
INTRODUCCION
Desde sus primeros inicios el hombre ha seleccionado plantas y domesticado animales mediante el cruzamiento selectivo de individuos con el fin de transferir los caracteres deseados. La principal limitante de este proceso radica en la incompatibilidad sexual observada cuando los organismos son muy divergentes genéticamente, lo que impide esta transferencia entre especies. La ingeniería genética permite romper esta barrera posibilitando la incorporación de genes desde otras especies que de otra forma sería imposible con los métodos de mejoramiento tradicional. De esta forma los organismos genéticamente modificados o más comúnmente denominados transgénicos son organismos vivos (plantas, animales o bacterias) manipulados genéticamente mediante la inserción de un gen que habitualmente no formaba parte de su repertorio genético. La finalidad de esto es proporcionar a la planta o animal nuevas características productivas y hacerlos más eficientes y competitivos (Clark, 2002).
El gen a ser introducido consiste básicamente de una construcción de ADN que contiene una región promotora y la región codificante para la proteína de interés, los que se insertan en el genoma en forma artificial mediante alguna de las técnicas disponibles como se detalla más adelante. Esta región de ADN es denominada comúnmente transgen y puede provenir de otro animal de la misma especie o desde una bacteria o planta. El lugar del animal donde se expresa el transgen está dirigido por la región promotora o las regiones reguladoras de la región promotora. Así por ejemplo, si se utiliza el promotor del gen de la ß-Lactoglobulina se podría dirigir la expresión y secreción de una proteína recombinante exclusivamente en la leche del animal.
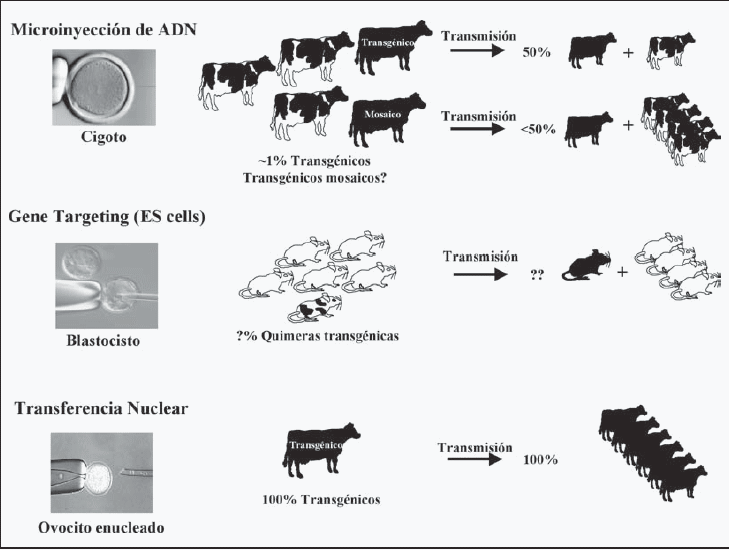
Hasta muy recientemente, la tecnología para generar animales de granja modificados genéticamente involucraba la microinyección pronuclear, en la cual pequeñas cantidades del ADN de interés (transgen) eran inyectadas en el pronúcleo de un embrión al estado de dos células. Aunque ampliamente aceptada y utilizada en forma rutinaria en muchos laboratorios, ha habido muy poco progreso para mejorar su eficiencia, la que se mantiene en el orden 0.1-5%, dependiendo de la especie considerada (Wall y col., 1997). Al mismo tiempo que la microinyección pronuclear se estaba desarrollando, en animales de granja, las células madre embrionarias (ES cells) estaban siendo desarrolladas y utilizadas para experimentos de recombinación homóloga en el ratón. Esta tecnología ha sido tremendamente eficiente para explotar la capacidad de las células en contribuir a la línea germinal y realizar recombinación homóloga con ADN exógeno, permitiendo la introducción de cambios precisos en el genoma del animal (para revisión ver Hooper, 1992). Sin embargo, a pesar de los esfuerzos de muchos laboratorios, no se han descrito aún células madre embrionarias en otras especies distintas al ratón (Stice, 1998), lo que ha frenado muchas aplicaciones potenciales de esta tecnología en animales de granja. Esta situación se revirtió recientemente después del nacimiento de Dolly, el primer animal obtenido por transferencia nuclear de una célula adulta (Wilmut y col., 1997). Sin embargo Polly, la primera oveja transgénica obtenida por transferencia nuclear (Schnieke y col., 1997), y George y Charlie, los primeros terneros transgénicos obtenidos de la misma forma (Cibelli y col., 1998), son los que han marcado el curso de las investigaciones en este campo y conducirán el camino en el futuro. Hoy en día, la transferencia nuclear se ha convertido en una alternativa real a la microinyección pronuclear debido a que permite un acortamiento del tiempo entre la obtención de un animal transgénico fundador y el establecimiento de un rebaño transgénico. Además, la transferencia nuclear abrió las puertas a la recombinación homóloga de animales de granja que había sido restringida sólo al ratón mediante la tecnología de células madre embrionarias. Esto posibilita la eliminación de genes endógenos en animales de granja (gene knock out) y/o la inserción de genes específicos en regiones transcripcionalmente activas del genoma, favoreciendo altos niveles de expresión de la proteína de interés.
METODOS DE TRANSFORMACION GENETICA EN ANIMALES
Transformación genética mediante el uso de vectores retrovirales. Contrario a la percepción de muchos, los primeros animales transgénicos fueron producidos hace ya casi 30 años mediante la microinyección de ADN viral (SV40) en la cavidad del blastocele de embriones de ratón (Jaenish y Mintz, 1974). Los próximos intentos involucraron embriones de ratón infectados con el retrovirus Moloney de la leucemia murina (MoMuLV), lo que resultó en la transmisión estable hacia la línea germinal (Jaenish, 1976). Esto se logró reemplazando genes que no son esenciales para el virus por genes heterólogos, aprovechando así la capacidad de los virus de infectar un amplio espectro de células y con una gran eficiencia. Una de las grandes desventajas de este método radica en que la integración del ADN se produce en diferentes etapas del embrión en desarrollo, lo que implica que el ADN no se integra en todas las células somáticas o en la línea germinal y por lo tanto no hay transmisión del transgen a la descendencia. Además, los animales generados por este método tienen a menudo más de un sitio de integración, lo cual ocurre cuando más de una célula del embrión es infectada por el virus (Palmiter y Brinster, 1985). Esto implica que las líneas de ratones transgénicos deben ser cruzadas para segregar los diferentes loci conteniendo el transgen y poder así aislar líneas con un sitio de inserción único. Finalmente, los vectores retrovirales poseen una limitada capacidad de ADN foráneo que puede ser acomodado, alrededor de 8 kb, lo cual imposibilita muchos experimentos especialmente con secuencias genómicas humanas que pueden superar ampliamente este tamaño.
Transformación genética mediante microinyección pronuclear. En la década del 80 ocurrió un importante avance en la tecnología de animales transgénicos que marcó el curso de la investigación en este campo por al menos dos décadas. Gordon y colaboradores describieron una técnica donde el ADN desnudo fue inyectado en el pronúcleo de un ovocito de ratón recientemente fertilizado, el que posteriormente se transfirió a hembras receptoras sincronizadas (Gordon y col., 1981). Este experimento demostró que era posible usar un plásmido recombinante como vector para transferir genes foráneos directamente hacia el embrión. El ADN inyectado de esta forma se integró en el genoma y pudo ser heredado por la descendencia de los animales transgénicos fundadores. La inyección de embriones al estado de una célula fue clave para obtener una integración temprana del transgen, permitiendo al ADN foráneo contribuir en el genoma de todas las células somáticas y la línea germinal. En ciertos casos la integración del transgen también ocurrió después de la primera división del zigoto, lo que resultó en animales fundadores mosaicos. Estos animales mosaicos aún transmiten el transgen a la descendencia pero lo hacen a una frecuencia menor al 50%. Desgraciadamente, la eficiencia para generar animales transgénicos utilizando esta tecnología es baja, particularmente en animales de granja. La eficiencia de la inyección pronuclear está controlada por una serie de factores como quedara demostrado por los trabajos de Brinster y colaboradores, quienes entregaron valiosa información sobre la integración de los transgenes y permitieron establecer que la concentración y la forma (circular o linear) del ADN eran los factores más críticos para una eficiente integración (Brinster y col., 1985). No se encontraron diferencias significativas cuando el pronúcleo femenino o masculino era utilizado para la inyección, aunque este último es preferido por ser más grande. Por otro lado, el cruzamiento de ratones híbridos (por ejemplo C57BL x SJL) fue más exitoso en la producción de animales transgénicos que las líneas cosanguíneas (C57Bl x C57Bl). El descubrimiento de la inyección de pronúcleos como un nuevo método para modificar el genoma de los animales revolucionó la forma en que los investigadores pudieron analizar la expresión de los transgenes y pavimentó el camino para la generación de los primeros animales transgénicos de granja hace ya 18 años (Hammer y col., 1985). Desde entonces, la tecnología ha sido implementada con éxito en la mayoría de los animales domésticos como en conejos (Buhler y col., 1990), ovejas (Wright y col., 1991), cabras (Ebert y col., 1991), vacas (Krimpenfort y col., 1991) y cerdos (Wall y col., 1991). Sin embargo, además de los problemas asociados con la integración de los transgenes hay ineficiencias asociadas con la recolección, cultivo de los huevos fertilizados y transferencia de los embriones hacia las hembras receptoras. Otros factores como el largo período de gestación y el bajo número de animales por generación, sumado al costo extra de cuidado de los animales, han contribuido a la lenta adopción de estas tecnologías, especialmente en los países menos desarrollados.
Los primeros estudios realizados en animales de granja se enfocaron hacia el uso de genes que controlan la productividad del animal, por ejemplo genes de la hormona del crecimiento para incrementar la tasa de crecimiento y la eficiencia de conversión (Pursel y Rexroad, 1993). Estos estudios mostraron los problemas de la inyección pronuclear con relación al control de los niveles de expresión del transgen. Se encontró una gran variación de expresión en las líneas de animales transgénicos generados, siendo ésta en general muy pobre, especialmente si no todos los elementos reguladores del transgen eran incluidos en la construcción genética (plásmido). Esto llevó a que muchos investigadores dedicaran mayores esfuerzos a entender la forma en que los transgenes se insertan en el genoma del animal y los factores que afectan la expresión de los mismos. Esto explica por qué la mayoría de los experimentos dirigidos a alterar la composición de la leche han sido realizados principalmente en el ratón, aunque existen algunas notables excepciones tales como la secreción de proteínas de valor terapéutico como el factor IX de la coagulación en la leche de ovejas (Wright y col., 1991), el activador de plasminógeno de tejido en la leche de cabras (Ebert y col., 1991) y la lisozima humana en la leche de bovinos (Krimpenfort y col., 1991). Sin embargo, dada la baja eficiencia de esta tecnología (Clark y col., 1994), sumado a los costos involucrados, es que ha habido una búsqueda constante por nuevas alternativas. Así, la manipulación de las células madre embrionarias de ratón (ES cells) apareció como una potencial solución para muchos de los problemas encontrados con la técnica de microinyección pronuclear.
Transformación genética mediante recombinación homóloga en células madre embrionarias (ES cells). El aislamiento de células madre embrionarias de ratón, en 1989, abrió nuevas posibilidades para estudiar la función génica en animales transgénicos (Thompson y col., 1989). Las células madre embrionarias se obtienen desde el macizo celular interno de blastocistos y se pueden mantener en cultivos sin perder su estado indiferenciado gracias a la presencia en el medio de cultivo de factores inhibitorios de la diferenciación. Estas células pueden ser manipuladas in vitro vía recombinación homóloga, permitiendo así alterar la función de genes endógenos. Las células así modificadas pueden ser entonces reintroducidas en blastocistos receptores contribuyendo eficientemente a la formación de todos los tejidos en un animal quimérico incluyendo la línea germinal (Evans y Kaufman, 1981; Martin, 1981). Esta tecnología posibilita modificaciones genéticas muy finas en el genoma del animal, tal como la introducción de copias únicas de un gen. La incorporación de una copia única de un gen en un sitio predeterminado del cromosoma tiene las ventajas de permitir controlar el número de copias del transgen y se puede controlar la inserción de este en un sitio favorable (transcripcionalmente activo) para su expresión tejido-específica. Utilizando esta tecnología ha sido posible anular la función de genes endógenos del ratón mediante la integración de un marcador de selección, lo que ha permitido la generación de varios cientos de ratones llamados knock out que han servido como modelos de enfermedades genéticas en humanos y como modelos para analizar la función de genes endógenos (Melton, 1994; Shastry, 1998; Kolb y col., 1999; Wallace y col., 2000). Lamentablemente, la generación de animales modificados genéticamente a través de esta ruta ha sido limitada sólo al ratón, fundamentalmente por la imposibilidad de aislar células madre embrionarias de otras especies que conserven la capacidad de totipotencialidad que caracteriza a estas células (Clark y col., 1992; Niemann y Reichelt, 1993; Cibelli y col., 1998). Aunque células parecidas a las células madre embrionarias (ES-like cells) han sido descritas en otras especies (lo que ha contribuido a la formación de vacas y cerdos quiméricos a partir de ellas), en ningún caso se ha demostrado que puedan transmitir las modificaciones a su descendencia, lo cual ha impedido el establecimiento de líneas de animales transgénicos a través de esta ruta (Niemann y Reichelt, 1993; Cibelli y col., 1998).
Transformación genética mediada por semen. En 1989 Lavitrano y col. describieron la producción de ratones transgénicos mediante inseminación artificial, utilizando semen que había sido incubado con ADN exógeno. Aunque atractivo por su simplicidad, este procedimiento ha sido muy cuestionado por su poca reproducibilidad, ya que a pesar de los esfuerzos de los principales laboratorios del mundo estos experimentos no pudieron ser repetidos en el ratón (Brinster y col., 1989). Más recientemente, el mismo grupo clama haber producido cerdos transgénicos con una muy alta eficiencia de 54-60% (Lavitrano y col., 1997); sin embargo, la integridad del transgen no fue analizada y otros estudios demuestran que el ADN internalizado por esta ruta sufre re-arreglos y recombinación con el ADN genómico, lo que dificulta la formación de líneas de animales transgénicos con niveles de expresión estable (Zoraqi y Spadafora, 1997).
Transformación genética mediante transformación de células somáticas y transferencia nuclear (clonación). La transferencia nuclear se describió por primera vez en 1952 en anfibios y consiste en extraer el material genético de un ovocito para posteriormente introducirle el material genético de una célula del animal a clonar. Los trabajos pioneros de Briggs y King (1957) demostraron que los núcleos de células al estado de blastocisto eran capaces de dirigir el desarrollo embrionario normal de ovocitos reconstituidos y generar una rana adulta a partir de estas células. En la década de los ochenta se realizaron exitosamente transferencias nucleares en la mayoría de los mamíferos como conejos (Stice y Robl, 1988), cerdos (Prather, y col., 1989), bovinos (Prather y col., 1987) y ovejas (Willadsen, 1986; Smith y Wilmut, 1989). Estos experimentos se realizaron por medio de la disociación de blastómeros embrionarias (células embrionarias no diferenciadas) y su posterior transferencia nuclear. Sin embargo, los intentos por realizar transferencia nuclear con células más diferenciadas fueron infructuosos, lo que llevó a pensar que el ADN de células diferenciadas no podía reprogramarse, surgiendo entonces el dogma de que el proceso de diferenciación celular era irreversible. Este dogma se derrumbó el 27 de febrero de 1997, fecha en que Ian Wilmut y sus colegas del Instituto Roslin, en Edimburgo, Escocia, explicaron en la revista Nature cómo habían creado a la oveja Dolly (figura 1). Dolly fue el resultado de la fusión de un núcleo procedente de una célula mamaria extraída de una oveja adulta con un óvulo al que previamente se le había extraído el material genético (proceso conocido como enucleación). El equipo escocés demostraba así que las células adultas y especializadas podían ser reprogramadas. La etapa clave de este proceso fue la coordinación del ciclo celular de la célula receptora y el de la célula donante de núcleos que se logró mediante la deprivación de suero de estas últimas (Campbell y col., 1996; Wilmut y col., 1997). La deprivación de suero, previo a la transferencia nuclear, induce a las células a salir del ciclo de crecimiento y entrar en un estado de arresto celular o quiescencia (G0/G1 en el ciclo celular), que se ha propuesto potenciaría el desarrollo embrionario, permitiendo a los factores en el ooplasma reprogramar el núcleo donante (Campbell y col., 1996). Siguiendo los hallazgos de Campbell y Wilmut muchos grupos han reproducido exitosamente estos experimentos (Schnieke y col., 1997; Wells y col., 1997; Baguisi y col., 1999; Zakhartchenko y col., 1999a; Reggio y col., 2001), con la excepción de un grupo en Estados Unidos, quienes han recomendado el uso de células proliferando activamente, esto es, células en G1 en el ciclo celular en vez de deprivadas de suero (Cibelli y col., 1998). Sin embargo, ninguno de estos estudios comparó directamente la eficiencia de ambos tratamientos para producir animales clonados y estudios recientes, utilizando fibroblastos fetales, han demostrado que la inducción del estado de quiescencia en las células mejora significativamente el desarrollo a la etapa de blastocisto comparado con células proliferativas, aunque no se evaluó la tasa de éxito de la transferencia de estos embriones en receptoras sincronizadas (Hill y col., 2000; Zakhartchenko y col., 1999b).
|
FIGURA 1. Dolly: El primer mamífero clonado por transferencia nuclear desde una célula adulta. |
Dolly: The first mammal ever cloned by nuclear transfer from an adult mammary cell. |
La clonación utilizando células somáticas sin transformar ha demostrado la utilidad en generar clones de animales individuales de alto mérito genético; sin embargo, el real potencial de la tecnología radica en la generación de animales transgénicos, lo que es posible mediante la incorporación de los genes de interés a las líneas celulares mediante transfección que puede ser realizada vía lipofección (Watanabe y col., 1994) o por electroporación (Wong y Neumann, 1982; Oshima y col., 1998). Las células transformadas, que se seleccionan previamente mediante la incorporación de genes de resistencia, pueden ser utilizadas como donantes de núcleos en la transferencia nuclear, obteniéndose de esta forma clones de animales que además son transgénicos (Schnieke y col.,1997; Cibelli y col., 1998; McCreath y col., 2000). A diferencia de la microinyección pronuclear, donde sólo un 3-5% de los animales nacidos son transgénicos (cifra que es inferior en animales mayores), la transferencia nuclear asegura que el 100% de los animales nacidos sea transgénico, eliminándose, por lo tanto, una generación de animales (figura 2). Además, estos animales presentan un bajo índice y/o ausencia total de mosaiquismo. Esto permite producir varios animales transgénicos en la primera generación, posibilitando hacer pruebas de expresión del transgen en un grupo de animales mientras el resto se utiliza para propagar la línea. En forma adicional es posible seleccionar el sexo del animal sin necesidad de incurrir a biopsias del embrión, lo que permitiría incrementar la masa ganadera por multiplicación (clonación) en forma más rápida y eficiente. Finalmente, la posibilidad de recombinación homóloga (gene targeting) en células somáticas previo a la transferencia nuclear abre infinitas posibilidades de manipulación genética en animales de granja que habían sido restringidas sólo al ratón, a través de recombinación homóloga en células madre embrionarias. Esto permitirá expandir el horizonte de posibilidades a esta tecnología, posibilitando la eliminación de genes de interés en el animal (gene knock out), como, por ejemplo, la eliminación del gen del cerdo responsable del rechazo a los trasplantes de órganos (Phelps y col., 2003; Ramsoondar y col., 2003) o la eliminación del gen de la oveja responsable de la producción de priones (Denning y col., 2001), además de la posibilidad de insertar genes específicos en regiones definidas del genoma del animal, favoreciendo un sitio permisivo para asegurar altos niveles de expresión de la proteína de interés (McCreath y col., 2000).
POTENCIALES USOS DE LA TECNOLOGIA DE TRANSFERENCIA NUCLEAR Y RECOMBINACION HOMOLOGA EN PRODUCCION ANIMAL
Clonación de animales elite. Además de proporcionar una ruta para la generación de animales transgénicos, la transferencia nuclear podría utilizarse para realizar la imagen más popular de la clonación, esto es, la producción de cantidades ilimitadas de animales genéticamente idénticos. La posibilidad de multiplicar razas de animales seleccionados podría aumentar la eficiencia de la productividad pecuaria. Sin embargo, la principal ventaja de la clonación no sería en los programas de selección, sino en la diseminación más rápida del progreso genético desde rebaños elite hacia los productores. Hasta la fecha esto se ha venido realizando mediante la inseminación artificial, la cual suministra sólo la mitad de los genes. Con la clonación, los productores que pudieran pagar este servicio recibirían embriones que serían clones de las vacas más productivas de los rebaños elite, con lo que incrementarían la performance de sus rebaños en tan sólo una generación. En este escenario las empresas venderían embriones clonados de la misma forma en que hoy comercializan el semen. Estos embriones tendrían la ventaja de un transporte más fácil de los genotipos entre países, evitándose los inconvenientes de la cuarentena. Un potencial riesgo de esta práctica estaría en la posibilidad de pérdida de diversidad genética; sin embargo, esto se podría evitar restringiendo la venta de un número limitado de clones de cada genotipo a cada productor. Aunque los rebaños de algunos productores pudieran consistir sólo de animales clonados, el hecho de que estos fueran clones de diferentes animales elite incrementaría la diversidad genética en estos predios.
Conservación genética. Aunque la transferencia nuclear se asocia en la mente de la gente con una pérdida de la diversidad genética, esta técnica también proporciona nuevas alternativas para la conservación genética. Con una cada vez más creciente presión comercial, muchas razas indígenas o criollas adaptadas a las condiciones locales (a modo de ejemplo la raza Overo Negro en nuestro país) están siendo reemplazadas por razas comerciales sujetas a sistemas intensivos de producción. Estas razas locales pueden contener importantes genes que confieran resistencia a enfermedades y resistencia a las condiciones climáticas (frío/calor). Hay, por tanto, una urgente necesidad por prevenir su extinción. Los métodos actuales de conservación consisten en almacenar semen o embriones congelados, procesos que son largos y costosos. Como consecuencia, el futuro de sólo unas pocas razas está asegurado. La tecnología de clonación puede proporcionar una forma más simple y efectiva de conservar estas razas, por cuanto muestras de sangre, biopsias de piel o incluso pelo pueden ser utilizadas como fuentes de células que podrían ser crecidas brevemente en el laboratorio, mantenidas y congeladas mediante su almacenamiento en nitrógeno líquido para ser luego utilizadas en experimentos de transferencia nuclear. El mejor ejemplo del potencial de esta tecnología lo demuestran los recientes experimentos en diversas especies en peligro de extinción mediante transferencia nuclear interespecies (Lanza y col., 2000; Kitiyanant y col., 2001; Loi y col., 2001; Lee y col., 2003).
Eliminación de genes (gene knock out). La posibilidad de recombinación homóloga (gene targeting) en células somáticas previo a la transferencia nuclear ha captado la atención de las principales compañías biotecnológicas que desean capitalizar los frutos de esta tecnología (Pollock y col., 1999; Reggio y col., 2001). La modificación genética que conduce a la pérdida de función de un gen, o más comúnmente conocida como gene knock out, ha sido utilizada extensivamente en el ratón para obtener un mayor entendimiento de la función génica y como modelo para ciertas enfermedades (Shastry, 1998; Kolb y col., 1999; Wallace y col., 2000). Actualmente existe una gran carencia de órganos para trasplantes humanos que no es cubierta por las donaciones. El trasplante de órganos de animales hacia humanos (xenotrasplantes) podría ser la solución. Sin embargo, existen muchas barreras de rechazo entre especies que limitan su uso. La principal barrera consiste en el fenómeno de rechazo hiperagudo debido a la presencia de anticuerpos naturales contra epítopes del disacárido galactosa 1-3 galactosa presente en las superficies celulares de mamíferos, pero que estarían ausentes en las superficies celulares de humanos y monos (Gallili y col., 1985). Este es uno de los principales usos donde la tecnología de recombinación homóloga acoplada con transferencia nuclear está siendo explotada. La reciente generación de cerdos transgénicos en los que se eliminó el gen 1-3 galactosiltransferasa y que permitiría la producción de animales que carecen del epítope responsable del rechazo hiperagudo, es una clara demostración del poder de esta tecnología (Phelps y col., 2003).
Otra de las aplicaciones de esta tecnología está en la creación de animales resistentes a ciertas enfermedades. Las enfermedades ocasionadas por priones han tenido un enorme impacto económico en algunos países de Europa y la utilización de productos animales para su uso en humanos es una preocupación constante. Este es el caso de la encefalopatía espongiforme bovina (EEB) o más comúnmente conocida como enfermedad de las vacas locas que sería la causa de una nueva forma de enfermedad de Creuzfeldt Jacob en humanos denominada vCJD (Hill y col., 1997). Experimentos realizados en ratones (Prusiner y col., 1993) y más recientemente en ovejas (Denning y col., 2001), demuestran que es factible eliminar el gen para los priones (PrP) mediante recombinación homóloga y que los animales producidos son resistentes al Scrapie. Dado que las ovejas y vacas están siendo utilizadas para producir proteínas humanas de uso farmacológico y que estos animales poseen genes funcionales PrP, sería apropiado producir poblaciones de animales resistentes a estos priones.
La tecnología de recombinación homóloga ha sido ampliamente utilizada en el ratón para producir varios modelos de enfermedades humanas. Sin embargo, debido a las diferencias fisiológicas sería más apropiado disponer de modelos animales que se asemejen más al humano. Este es el caso del modelo para fibrosis quística creado en el ratón, donde se eliminó el gen Cftr (Cystic fibrosis transmembrane responder). Este modelo no presentó las características típicas de la enfermedad en humanos debido a las diferencias en la fisiología del pulmón del ratón (Davidson y col., 1995). La eliminación del gen Cftr en la oveja se esperaría produjera un modelo animal para fibrosis quística más exacto debido a las similitudes de la fisiología pulmonar entre ovejas y humanos (Harris, 1997).
Las modificaciones en los animales que pudieran influir rasgos de producción tales como el crecimiento y la eficiencia de alimentación son uno de los principales objetivos en el mejoramiento animal. Ratones en donde el gen de la miostatina se eliminó mediante recombinación homóloga desarrollaron mayor musculatura esquelética que los controles no modificados (McPherron y col., 1997). Además, el fenotipo de doble musculatura de algunas razas bovinas, por ejemplo Belgian Blue, ha sido asociado a modificaciones (mutaciones naturales) del gen de la miostatina (Grobet y col., 1997). Por lo tanto, la eliminación de este gen en bovinos, ovejas y cerdos podría producir animales con mayor masa muscular, lo cual sería de enorme importancia económica. Tal es así que actualmente existe una empresa biotecnológica en USA (http:// www.prolinia.com) cuyo objetivo es proporcionar la clonación a los mejoradores de razas registradas, para luego mediante ingeniería genética proveer a estas razas elite de características deseables como la mutación para el gen de la miostatina.
Otra característica productiva que se podría mejorar a través de esta tecnología es la leche. La leche aporta cerca del 30% de las proteínas consumidas en los países desarrollados. Por esta razón, la lactancia ha sido objeto de diversos estudios en el campo de genética, fisiología y nutrición. La recombinación homóloga podría ser utilizada para reemplazar genes de las proteínas de la leche de los animales de granja con la respectiva contraparte de los genes humanos para utilizarlos como fuentes de proteínas. Por ejemplo, la seroalbúmina humana es utilizada ampliamente para el tratamiento de quemaduras y como reemplazo de fluidos corporales en cirugía. La escala de requerimiento de esta proteína (~ 600 toneladas al año) la hacen un muy buen candidato para su producción a escala comercial en la leche de vacas transgénicas. Desafortunadamente, la seroalbúmina bovina es muy similar a la humana, lo que genera problemas para su purificación. Una solución sería reemplazar el gen bovino con su contraparte humana, así la proteína bovina sería eliminada sin alterar o comprometer la viabilidad del animal. Otro ejemplo lo constituye la ß-Lactoglobulina que está presente sólo en la leche de rumiantes y no tiene una función conocida en el proceso de secreción de leche (Clark y col., 1998). Su presencia confiere algunas propiedades de elaboración indeseadas y se cree es la responsable de la mayoría de las alergias a la leche bovina, situación que afecta a una considerable parte de la población mundial. Por lo tanto, la eliminación de esta proteína podría proporcionar nuevas propiedades tecnológicas a la leche (Richardson, 1985). Además, su eliminación no sólo ayudaría con el problema de las alergias, sino que también, debido a la compensación compensación que se produciría en la concentración de las otras proteínas de la leche, probablemente incrementaría la concentración de caseínas, lo que tendría un efecto directo para la industria quesera. De la misma forma, la sobreexpresión de caseínas en la leche se esperaría que alterara significativamente las propiedades tecnológicas de la misma como fuera demostrado recientemente por Brophy y col., (2003) y la expresión de proteasas podría generar resistencia a enfermedades de gran impacto en el sector lechero como la mastitis (Kerr y col., 2001).
CONCLUSION
La combinación de las tecnologías de transferencia nuclear y recombinación homóloga hacen posible actualmente realizar modificaciones genéticas muy precisas en el genoma del animal y han contribuido a incrementar la eficiencia del proceso de generación de animales transgénicos de granja. Aunque en una primera etapa las modificaciones genéticas afectarán solamente a genes simples, a medida que la tecnología avance y se haga más eficiente es posible vislumbrar un mayor rango de modificaciones que irán desde cambios en unos pocos pares de bases hasta la reingeniería de grandes segmentos cromosómicos. Muchas aplicaciones requerirán de modificaciones genéticas múltiples, lo que puede significar un problema para los animales de granja dado su largo período generacional. Por lo tanto, el real desafío será desarrollar células que mantengan su capacidad de totipotencialidad para transferencia nuclear después de múltiples eventos de recombinación homóloga. Esto permitirá realizar más de una modificación genética en el animal y acortará el tiempo necesario para establecer estos cambios al estado de homocigosis. Aunque por el momento muchas de las aplicaciones de esta tecnología están orientadas principalmente a la industria farmacéutica, es posible vislumbrar que rasgos tan complejos de manipular como una mayor eficiencia en la conversión de alimentos y la resistencia a enfermedades, entre otras, serán más fácilmente abordadas gracias a esta tecnología.
BIBLIOGRAFIA
BAGUISI, A., E. BEHBOODI, D. MELICAN, J. POLLOCK, M. DESTREMPES, C. CAMMUSO, J. WILLIAMS, S. NIMS, C. PORTER, P. MIDURA, M. PALACIOS, S. AYRES, R. DENNISTON, M. HAYES, C. ZIOMEK, H. MEADE, R. GODKE, W. GAVIN, E. OVERSTROM, Y. ECHELARD. 1999. Production of goats by somatic cell nuclear transfer. Nat Biotechnol. 17: 456-61.
BRIGGS, R., T. KING. 1957. Changes in the nuclei of differentiating endodermal cells as revelead by nuclear transplantation. J. Morphol. 100: 269-312.
BRINSTER, R., H. CHEN, M. TRUMBAUER, M. YAGLE, R. PALMITER. 1985. Factors affecting the efficiency of introducing foreign DNA into mice by microinjecting eggs. Proc Natl Acad Sci. U S A. 82: 4438-42.
BRINSTER, R., E. SANDGREN, R. BEHRINGER, R. PALMITER. 1989. Non simple solution for making transgenic mice. Cell. 59: 239-41.
BROPHY, B., G. SMOLENSKI, T. WHEELER, D. WELLS, P. L'HUILLIER, G. LAIBLE. 2003. Cloned transgenic cattle produce milk with higher levels of beta-casein and kappa-casein. Nat Biotechnol. 21: 157-62.
BUHLER, T., T. BRUYERE, D. WENT, G. STRANZINGER, K. BURKI. 1990. Rabbit beta-casein promoter directs secretion of human interleukin-2 into the milk of transgenic rabbits. Biotechnology. 8: 140-3.
CAMPBELL, K., J. MCWHIR, W. RITCHIE, I. WILMUT. 1996. Sheep cloned by nuclear transfer from a cultured cell line. Nature 7: 64-6.
CIBELLI, J., S. STICE, P. GOLUEKE, J. KANE, J. JERRY, C. BLACKWELL, F. PONCE DE LEON, J. ROBL. 1998. Cloned transgenic calves produced from nonquiescent fetal fibroblasts. Science 22: 1256-8.
CLARK, A.J., A. COWPER, R. WALLACE, G. WRIGHT, J. SIMONS. 1992. Rescuing transgene expression by co-integration. Biotechnology. 10: 450-4.
CLARK, A.J., P. BISSINGER, D. BULLOCK, S. DAMAK, R. WALLACE, C. WHITELAW, F. YULL. 1994. Chromosomal position effects and the modulation of transgene expression. Reprod Fertil Dev. 6: 589-98.
CLARK, A., S. ALI, A. ARCHIBALD, H. BESSOS, P. BROWN, S. HARRIS, M. McCLENAGHAN, C. PROWSE, J. SIMONS, B. WHITELAW. 1998. The molecular manipulation of milk composition. Genome. 31: 950-5.
CLARK, A. 2002. Generation of transgenic livestock by pronuclear injection. Methods Mol. Biol. 180: 273-87.
DAVIDSON, D., J. DORIN, G. MCLACHLAN, V. RANALDI, D. LAMB, C. DOHERTY, J. GOVAN, D. PORTEOUS. 1995. Lung disease in the cystic fibrosis mouse exposed to bacterial pathogens. Nat. Genet. 9: 351-7.
DENNING, C., P. DICKINSON, S. BURL, D. WYLIE, J. FLETCHER, AJ. CLARK. 2001. Gene targeting in primary fetal fibroblasts from sheep and pig. Cloning Stem Cells 3: 221-31.
EBERT, K., J. SELGRATH, P. DITULLIO, J. DENMAN, T. SMITH, M. MEMON, J. SCHINDLER, G. MONASTERSKY, J. VITALE, K. GORDON. 1991. Transgenic production of a variant of human tissue-type plasminogen activator in goat milk: generation of transgenic goats and analysis of expression. Biotechnology. 9: 835-8.
EVANS, M., M. KAUFMAN. 1981. Establishment in culture of pluripotential cells from mouse embryos. Nature. 9: 154-6.
GALLILI, U., B. MACHER, J. BUEHLER, S. SHOHET. 1985. Human natural anti-alpha-galactosyl IgG. II. The specific recognition of alpha (1-3)-linked galactose residues. J. Exp. Med. 162: 573.
GORDON, J., F. RUDDLE. 1981. Integration and stable germ line transmission of genes injected into mouse pronuclei. Science 214: 1244-6.
GROBET, L., L. MARTIN, D. PONCELET, D. PIROTTIN, B. BROUWERS, J. RIQUET, A. SCHOEBERLEIN, S. DUNNER, F. MENISSIER, J. MASSABANDA, R. FRIES, R. HANSET, M. GEORGES. 1997. A deletion in the bovine myostatin gene causes the doublemuscled phenotype in cattle. Nat. Genet. 17: 71-4.
HAMMER, R., R. BRINSTER, R. PALMITER. 1985. Use of gene transfer to increase animal growth. Cold Spring Harb. Symp. Quant. Biol. 50: 379-87.
HARRIS, A. 1997. Towards an ovine model of cystic fibrosis. Hum. Mol. Genet. 6: 2191-4.
HILL, A., M. DESBRUSLAIS, S. JOINER, K. SIDLE, I. GOWLAND, J. DOEY, P. LANTOS. 1997. The same prion strain causes vCJD and BSE. Nature 389: 448-450.
HILL, J., Q. WINGER, C. LONG, C. LOONEY, J. THOMPSON, M. WESTHUSIN. 2000. Development rates of male bovine nuclear transfer embryos derived from adult and fetal cells. Biol. Reprod. 62: 1135-40.
HOOPER, M. 1992. Embryonal stem cells: introducing planned changes into the germline. H.J. Evans, ed. Harwood Academic Publishers, Switzerland. pp. 117.
JAENISH R., B. MINTZ. 1974. Simian virus 40 DNA sequences in healthy adult mice derived from preimplantation blastocist injected with viral DNA. Proc. Natl. Acad. Sci. USA 21: 1250-1254.
JAENISH, R. 1976. Germline integration and Mendelian transmission of the exogenous Molonkey leukaemia virus. Proc. Natl. Acad. Sci. USA 73: 1260.
KERR, D., K. PLAUT, A. BRAMLEY, C. WILLIAMSON, A. LAX, K. MOORE, K. WELLS, R. WALL. 2001. Lysostaphin expression in mammary glands confers protection against staphylococcal infection in transgenic mice. Nat. Biotechnol 19: 66-70.
KITIYANANT, Y., J. SAIKHUN, B. CHAISALEE, K. WHITE, K. PAVASUTHIPAISIT. 2001. Somatic cell cloning in Buffalo (Bubalus bubalis): effects of interspecies cytoplasmic recipients and activation procedures. Cloning Stem Cells 3: 97-104.
KOLB, A., R. ANSELL, J. MCWHIR, S. SIDDELL. 1999. Insertion of a foreign gene into the betacasein locus by Cre-mediated site-specific recombination. Gene. 227: 21-31.
KRIMPENFORT, P., A. RADEMAKERS, W. EYESTONE, A. VAN DER SCHANS, S. VAN DEN BROEK, P. KOOIMAN, E. KOOTWIJK, G. PLATENBURG, F. PIEPER, R. STRIJKER. 1991. Generation of transgenic dairy cattle using in vitro embryo production. Biotechnology 9: 844-7.
LANZA, R., J. CIBELLI, F. DIAZ, C. MORAES, P. FARIN, C. FARIN, C. HAMMER, M. WEST, P. DAMIANI. 2000. Cloning of an endangered species (Bos gaurus) using interspecies nuclear transfer. Cloning. 2: 79-90.
LAVITRANO, M., A. CAMAIONI, V. FAZIO, S. DOLCI, M. FARACE, C. SPADAFORA. 1989. Sperm cells as vectors for introducing foreign DNA into eggs: genetic transformation of mice. Cell. 57: 717-23.
LAVITRANO, M., M. FORNI, V. VARZI, L. PUCCI, M. BACCI, C. DI STEFANO, D. FIORETTI, G. ZORAQI, B. MOIOLI, M. ROSSI, D. LAZZERESCHI, A. STOPPACCIARO, E. SEREN, D. ALFANI, R. CORTESINI, L. FRATI. 1997. Sperm-mediated gene transfer: production of pigs transgenic for a human regulator of complement activation. Transplant Proc. 29: 3508-9.
LEE, B., G. WIRTU, P. DAMIANI, E. POPE, B. DRESSER, W. HWANG, B. BAVISTER. 2003. Blastocyst development after intergeneric nuclear transfer of mountain bongo antelope somatic cells into bovine oocytes. Cloning Stem Cells. 5: 25-33.
LOI, P., G. PTAK, B. BARBONI, J. FULKA, P. CAPPAI, M. CLINTON. 2001. Genetic rescue of an endangered mammal by cross-species nuclear transfer using post-mortem somatic cells. Nat Biotechnol. 19: 962-4.
MCCREATH, K., J. HOWCROFT, K. CAMPBELL, A. COLMAN, A. SCHNIEKE, A. KIND. 2000. Production of gene-targeted sheep by nuclear transfer from cultured somatic cells. Nature. 405: 1066-9.
MCPHERRON, A., A. LAWLER, S. LEE. 1997. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature. 387: 83-90.
MARTIN, G. 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. U S A. 78: 7634-8.
MELTON, W. 1994. Gene targeting in the mouse. Bioassays. 16: 633-8.
NIEMANN, H., B. REICHELT. 1993. Manipulating early pig embryos. J Reprod Fertil Suppl. 48: 5-94.
OSHIMA, Y., T. SAKAMOTO, I. YAMANAKA, T. NISHI, T. ISHIBASHI, H. INOMATA. 1998. Targeted gene transfer to corneal endothelium in vivo by electric pulse. Gene Ther. 5: 1347-54.
PALMITER, R., R. BRINSTER. 1985. Transgenic mice. Cell. 41: 343-5.
PHELPS, C., C. KOIKE, T. VAUGHT, J. BOONE, K. WELLS, S. CHEN, S. BALL, S. SPECHT, I. POLEJAEVA, J. MONAHAN, P. JOBST, S. SHARMA, A. LAMBORN, A. GARST, M. MOORE, A. DEMETRIS, W. RUDERT, R. BOTTINO, S. BERTERA, M. TRUCCO, T. STARZL, Y. DAI, D. AYARES. 2003. Production of alpha 1,3-galactosyltransferase-deficient pigs. Science 299: 411-4.
POLLOCK, D., J. KUTZKO, E. BIRCK-WILSON, J. WILLIAMS, Y. ECHELARD, H. MEADE. 1999. Transgenic milk as a method for the production of recombinant antibodies. J. Immunol. Methods. 231: 147-57.
PRATHER, R., F. BARNES, M. SIMS, J. ROBL, W. EYESTONE, N. FIRST. 1987. Nuclear transplantation in the bovine embryo: assessment of donor nuclei and recipient oocyte. Biol. Reprod. 4: 859-66.
PRATHER, R., M. SIMS, N. FIRST. 1989. Nuclear transplantation in early pig embryos. Biol. Reprod. 41: 414-8.
PRUSINER, S., D. GROTH, A. SERBAN, R. KOEHLER, D. FOSTER, M. TORCHIA, D. BURTON, S. YANG, S. DEARMOND. 1993. Ablation of the prion protein (PrP) gene in mice prevents scrapie and facilitates production of anti- PrP antibodies. Proc. Natl. Acad. Sci. U S A. 90: 10608-12.
PURSEL, V., C. REXROAD. 1993. Recent progress in the transgenic modification of swine and sheep. Mol. Reprod. Dev. 36: 251-4.
RAMSOONDAR, J., Z. MACHATY, C. COSTA, B. WILLIAMS, W. FODOR, K. BONDIOLI. 2003. Production of alpha 1,3-galactosyltransferase- knockout cloned pigs expressing human alpha 1,2-fucosylosyltransferase. Biol. Reprod. 69: 437-45.
REGGIO, B., A. JAMES, H. GREEN, W. GAVIN, E. BEHBOODI, Y. ECHELARD, R. GODKE. 2001. Cloned transgenic offspring resulting from somatic cell nuclear transfer in the goat: oocytes derived from both follicle-stimulating hormonestimulated and nonstimulated abattoir-derived ovaries. Biol. Reprod. 65: 1528-33.
RICHARDSON, T. 1985. Chemical modifications and genetic engineering of food proteins. J. Dairy Sci. 68: 2753-62.
SHASTRY, B.S. 1998. Gene disruption in mice: models of development and disease. Mol. Cell. Biochem. 1: 163-79.
SCHNIEKE, A., A. KIND, W. RITCHIE, K. MYCOCK, A. SCOTT, M. RITCHIE, I. WILMUT, A. COLMAN, K. CAMPBELL. 1997. Human factor IX transgenic sheep produced by transfer of nuclei from transfected fetal fibroblasts. Science 19: 2130-3.
SMITH, L., I. WILMUT. 1989. Influence of nuclear and cytoplasmic activity on the development in vivo of sheep embryos after nuclear transplantation. Biol. Reprod. 40: 1027-35.
STICE, S., J. ROBL. 1988. Nuclear reprogramming in nuclear transplant rabbit embryos. Biol Reprod. 39: 657-64.
STICE, S. 1998. Opportunities and challenges in domestic animal embryonic stem cell research. In: Animal Breeding: Technology for the 21st Century. AJ. Clark, ed. Harwood Academic Press, Switzerland, pp 64-71.
THOMPSON, S., A. CLARKE, A. POW, M. HOOPER, D. MELTON. 1989. Germ line transmission and expression of a corrected HPRT gene produced by gene targeting in embryonic stem cells. Cell 56: 313-21.
WALL, R., V. PURSEL, A. SHAMAY, R. MCKNIGHT, C. PITTIUS, L. HENNIGHAUSEN. 1991. High-level synthesis of a heterologous milk protein in the mammary glands of transgenic swine. Proc. Natl. Acad. Sci. U S A 88: 1696-700.
WALL, R., D. KERR, K. BONDIOLI. 1997. Transgenic dairy cattle: genetic engineering on a large scale. J. Dairy Sci. 80: 2213-2224.
WALLACE, H., R. ANSELL, J. CLARK, J. MCWHIR. 2000. Pre-selection of integration sites imparts repeatable transgene expression. Nucleic. Acids. Res. 28: 1455-64.
WATANABE, Y., H. NOMOTO, R. TAKEZAWA, N. MIYOSHI, T. AKAIKE. 1994. Highly efficient transfection into primary cultured mouse hepatocytes by use of cation-liposomes: an application for immunization. J. Biochem. 116: 1220-6.
WELLS, D., P.M. MISICA, T. DAY, H. TERVIT. 1997. Production of cloned lambs from an established embryonic cell line: a comparison between in vivo- and in vitro-matured cytoplasts. Biol. Reprod. 57: 385-93.
WILLADSEN, S.M. 1986. Nuclear transplantation in sheep embryos. Nature. 320: 63-5.
WILMUT, I., A. SCHNIEKE, J. MCWHIR, A. KIND, K. CAMPBELL. 1997. Viable offspring derived from fetal and adult mammalian cells. Nature 385: 810-3.
WONG T., E. NEUMANN. 1982. Electric field mediated gene transfer. Biochem. Biophys. Res. Commun. 30: 584-7.
WRIGHT, G., A. CARVER, D. COTTOM, D. REEVES, A. SCOTT, P. SIMONS, I. WILMUT, I. GARNER, A. COLMAN. 1991. High level expression of active human alpha-1- antitrypsin in the milk of transgenic sheep. Biotechnology 9: 830-4.
ZAKHARTCHENKO, V., R. ALBERIO, M. STOJKOVIC, K. PRELLE, W. SCHERNTHANER, P. STOJKOVIC, H. WENIGERKIND, R. WANKE, M. DUCHLER, R. STEINBORN, M. MUELLER, G. BREM, E. WOLF. 1999a. Adult cloning in cattle: potential of nuclei from a permanent cell line and from primary cultures. Mol. Reprod. Dev. 54: 264-72.
ZAKHARTCHENKO, V., G. DURCOVA-HILLS, M. STOJKOVIC, W. SCHERNTHANER, K. PRELLE, R. STEINBORN, M. MULLER, G. BREM, E. WOLF. 1999b. Effects of serum starvation and re-cloning on the efficiency of nuclear transfer using bovine fetal fibroblasts. J. Reprod. Fertil. 115: 325-31.
ZORAQI, G., C. SPADAFORA. 1997. Integration of foreign DNA sequences into mouse sperm genome. DNA Cell Biol. 16: 291-300.
Aceptado: 29-06-2004.