Arch. Med. Vet., Vol. XXXVI N° 2,
2004, pp. 155-162
ARTICULOS ORIGINALES
Concentraciones de inmunoglobulinas
séricas y calostrales de vacas selenio-deficientes y en el suero sanguíneo de
sus terneros
Serum and colostrum immunoglobulin
concentrations from selenium deficient cows and in the blood of their calves
V. LEYAN 1*,
M.V., M.Sc; F. WITTWER 2, M.V., M.Sc; P.A. CONTRERAS
2, M.V., M.Phil; J. KRUZE 3,
M.V., Ph.D.
1
Instituto de Inmunología, Facultad de Medicina, Universidad Austral de Chile,
Valdivia, Chile.
2 Instituto de Ciencias Clínicas Veterinarias, Universidad
Austral de Chile, Valdivia, Chile.
3 Instituto de Microbiología, Universidad Austral
de Chile, Valdivia, Chile.
Financiado por proyectos: DID-UACH 2002-60 y FONDECYT 119-993
*Correspondencia: e-mail: vleyan@uach.cl
Resumen
Se estudió el efecto de una dieta
selenio-deficiente en vacas gestantes sobre la concentración de inmunoglobulinas
en el calostro y en el suero sanguíneo de las crías en sus primeras horas de
vida. Para ello se trabajó con 12 vacas gestantes (7-8 meses) entre 4-9 años
de edad, mantenidas en estabulación permanente y sometidas a una dieta deficiente
en selenio (0.04 ppm de materia seca), constituida por heno y un alimento concentrado
comercial seleniodeficiente. Un grupo de seis vacas fueron suplementadas con
selenato de bario (1 mg Se/kg, s.c.). El balance de selenio fue medido a través
de la actividad de GSH-Px eritrocítica. La medición de inmunoglobulinas en el
suero y calostro de las vacas y en el suero de los terneros se realizó por inmunodifusión
radial.
Los resultados demostraron que no existen diferencias (P < 0.05) en la concentración
de IgG, IgM e IgA entre vacas suplementadas y no suplementadas. La deficiencia
de Se no afectó la concentración de IgG total, IgM e IgA en calostro; sin embargo
la suplementación con selenio disminuyó (P < 0.05) la concentración de IgG1.
Por otra parte, la deficiencia nutricional de Se en las vacas no afecta la concentración
de IgG total, IgG1, IgM e IgA en el suero de terneros en sus primeros seis días
de edad.
Palabras clave:
selenio, calostro, terneros, inmunoglobulinas.
Summary
The colostrum immunoglobulin
concentrations of selenium-deficient cows and in the blood of their calves were
studied in twelve Friesian cows of 4-9 year old. The animals were 7 months pregnant
at the beginning of the experience and it were fed with a selenium deficient
diet consisted in hay and a commercial concentrate (diet 0.04 ppm dry matter)
and kept under permanent housing condition with water ad libitum. The cows were
allocated into two homogeneous groups of six animals each, selenium supplemented
(Se-S) and selenium deficient (Se-D) and the animals of the Se-S group were
supplemented with barium selenate (1 mg/Se/kg s. c.) ± 45 days before calving,
whilst the Se-D group received nothing.
The balance of selenium was assessed by GSH-Px activity in blood and the concentration
of immunoglobulin in serum and colostrums by immunodiffusion radial method.
Selenium deficiency did not affect serum and colostrum total IgG, IgM and IgA
concentration in cows (P > 0.05). However, the IgG1 colostrum concentrations
decreased in selenium supplemented cows (P < 0.05). On the other hand, the nutritional
deficiency of selenium did not affect total IgG, IgG1, IgM and IgA in calves
serum concentrations until 6 days of age.
Key words:
selenium, colostrum, calves, immunoglobulins.
INTRODUCCION
El selenio (Se) juega un rol
importante en la integridad funcional del tracto reproductivo, la función tiroidea
y el normal funcionamiento del sistema inmunológico del ganado (López
Alonso y col., 1997). La principal fuente de Se para los bovinos a pastoreo
proviene de la alimentación con praderas naturales y forrajes conservados, que
en el Sur de Chile, en un alto porcentaje, son deficientes en este microelemento
(Ceballos y col., 1998). Para compensar la deficiencia nutricional
de Se se utiliza la suplementación con este mineral, permitiendo optimizar con
ello las variables productivas, de fertilidad y otorgar mayor protección a los
animales contra las enfermedades infecciosas (Wittwer y col.,
2002). En este sentido, en los últimos años hemos estudiado el efecto de
la deficiencia nutricional de Se en vacas lecheras estabuladas y en vaquillas
en pastoreo, demostrando sus efectos sobre la función tiroidea (Contreras
y col., 2002; Matamoros y col., 2003), la glándula mamaria
(Kruze y col., 2000) y el sistema inmune (Pesutic
y col., 2001). En esta última investigación se observó que la suplementación
con Se en vaquillas a pastoreo no favorece la respuesta inmune humoral ni celular.
Con relación al efecto del Se
sobre la concentración de inmunoglobulinas en el calostro y la absorción de
ellas en el ternero recién nacido, los estudios realizados son escasos y presentan
resultados variables. Por una parte, se ha informado que la administración de
Se solo o con vitamina E en vacas de carne, a mitad de preñez y pastoreando
en praderas selenio-deficientes, aumenta las concentraciones de IgG en el calostro
y en el suero de los terneros (Swecker y col., 1995). Sin
embargo, otros estudios señalan que el Se y la vitamina E favorecen la producción
de calostro y leche pero no benefician la inmunidad pasiva (Lacetera
y col., 1996).
La presente investigación tiene
por objetivo estudiar el efecto de la deficiencia nutricional de selenio en
vacas gestantes sobre la concentración de inmunoglobulinas IgG total, IgG1,
IgM e IgA en el calostro y en el suero sanguíneo de los terneros en sus primeras
días de vida.
MATERIAL Y METODOS
Animales.
Se utilizaron 12 vacas Frisonas de 4 a 9 años de edad (promedio de 5 años) con
7-8 meses de gestación, provenientes de un predio lechero libre de tuberculosis,
leucosis y brucelosis. Todos los animales fueron alimentados con una dieta deficiente
en Se (Se < 0.05 ppm) consistente en 9.5 kg de heno de pradera natural (Se=0.02
ppm de materia seca, MS), más 1 kg de concentrado comercial (Cosetan, Biomaster
IANSA-Chile) (Se=0.12 ppm de MS) y fueron mantenidos en estabulación permanente
durante todo el período experimental en cubículos individuales sobre piso de
hormigón y cama de paja, alimentados en comederos individuales y agua a voluntad.
Las vacas fueron distribuidas en dos grupos de seis animales, homogéneos en
términos de edad (6.5 años), número de partos (3.5 partos), peso corporal (594
kg), tiempo de gestación (230 días) y producción de leche en la última lactación
(6.406 litros). Uno de los grupos se suplementó con selenio (Se-S) y el otro
grupo no recibió suplementación (Se-D).
De las 12 vacas, una de ellas
perdió el ternero debido a un parto distócico. El examen post mortem del ternero
descartó la presencia de enfermedades infecciosas. De los 11 terneros nacidos
vivos, seis correspondieron a vacas Se-S y cinco al grupo Se-D. Las vacas permanecieron
tres días con sus terneros y posteriormente se les aplicó ordeño mecánico dos
veces al día. En el transcurso del período de estudio se registró la condición
corporal de todos los animales utilizando la escala 1 a 5 según Edmonson
y col. (1989).
Suplementación con
selenio y toma de muestras. Las
vacas del grupo Se-S fueron suplementadas por vía subcutánea con 1 mg de Se/kg
usando selenato de bario (Deposel®, Young Animal Health Ltd., N.
Zealand) en dosis única de 1 ml/ 50 kg/pv, administrado 60 días antes de la
fecha probable de los partos. De cada vaca se tomaron simultáneamente dos muestras
de sangre (con y sin heparina), por venopunción coccígea previo a la suplementación
(día 0) y luego cada 15 días durante 90 días. Inmediatamente después del parto
se recolectaron muestras de calostro de todas las vacas de ambos grupos y muestras
de suero de los terneros en la primera hora postnacimiento y cada dos días hasta
el sexto día de vida. Los sueros de las vacas, terneros y el calostro fueron
mantenidos a -20° C hasta su posterior análisis.
Evaluación de
la actividad sanguínea de glutatión
peroxidasa (GSH-Px). El balance nutricional de Se se evaluó mediante
la actividad sanguínea de GSH-Px realizado en un hemolizado de las muestras
de sangre con heparina de ambos grupos de vacas, empleando reactivos comerciales
(RANSEL® . Laboratorios Randox, Crumlin, UK), basados en una técnica
cinética compuesta NADPH-dependiente, según la modificación del método descrito
por Paglia y Valantine en 1967 (Ceballos y col., 1999). Las
lecturas se realizaron en un espectrofotómetro Hitachi 4020 (Roche Diagnostics
GmbH, Mannheim). La actividad de GSH-Px fue expresada en UI/g Hb e interpretada
según Ceballos y Wittwer (1996), quienes señalan que valores
menores a 60 UI/g Hb son considerados deficientes, entre 61 y 100 UI/g Hb son
considerados bajo marginales, 101 a 130 son marginales y valores superiores
a 130 UI/g Hb son considerados apropiados.
Determinación de
la concentración de inmunoglobulinas en
el suero y calostro. La determinación de
la concentración de inmunoglobulinas IgG total, IgG1, IgM e IgA en suero y calostro
de las vacas y suero de los terneros se realizó mediante un kit comercial de
inmunodifusión radial (BINDARID®. The Binding Site, UK). Previo a
la determinación se procedió a la separación de la grasa del calostro mediante
ultracentrifugación a 41.366 g por 1 hora, en una centrífuga refrigerada Sorvall
28S.
Análisis estadístico.
Establecida la normalidad de la distribución de los datos, mediante la prueba
de Kolmogorov y Smirnov, se calcularon los promedios y desviación estándar para
cada grupo y día de muestreo. La significancia de las diferencias dentro del
grupo se evaluó mediante análisis de varianza y cuando éstas fueron significativas
se identificaron con la prueba de comparación múltiple de Tukey. Las diferencias
entre los grupos se evaluaron mediante la prueba "t" de Student (Zar,
1996). Los análisis se realizaron con el programa Graph-Pad-Prism (versión
3), usándose un valor de significancia de P < 0.05.
RESULTADOS
La condición corporal de las vacas de ambos
grupos al comienzo del experimento fue de 3.3, disminuyendo durante el período
de estudio hasta 2.3, después del parto. No se presentaron diferencias entre
grupos (P > 0.05)
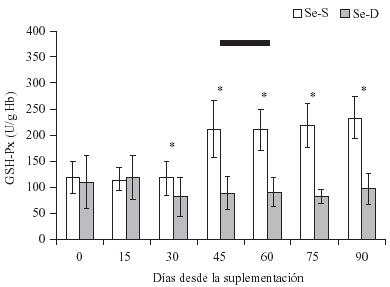
En el grupo Se-S se observó un
aumento (P < 0.05) de la actividad GSH-Px a partir del día 30 postsuplementación,
logrando valores considerados adecuados a los 45 días, efecto que se mantuvo
durante el período de estudio (figura 1); sin embargo, los
animales del grupo Se-D mantuvieron valores de actividad enzimática considerados
bajos (<100 U/g Hb), situación observada después de 30 días de consumo de la
ración selenio-deficiente, y se mantuvieron en esa condición durante el transcurso
del estudio, el que incluyó el período de los partos ocurridos entre los 45
a 60 días postsuplementación.
Durante el preparto, en ambos
grupos de animales se observó una disminución (P < 0.05) de la concentración
sérica de IgG total aproximadamente dos semanas antes del parto con respecto
a la sexta semana antes del parto (cuadro 1). La concentración
de IgG total, IgM e IgA en suero y calostro no mostró diferencias (P > 0.05)
entre los animales Se-S y Se-D. Sin embargo, la concentración de IgG1 fue mayor
(P < 0.05) en el calostro de las vacas no suplementadas con Se (cuadro
2).
Las concentraciones de inmunoglobulinas
séricas de los terneros de las vacas Se-S y Se-D aumentaron significativamente
(P < 0.05) entre la primera y las 48 horas de vida. Sin embargo, las concentraciones
de inmunoglobulinas IgG total, IgG1, IgM e IgA desde el nacimiento hasta el
sexto día de vida en los terneros de ambos grupos fueron similares (P < 0.05)
(cuadros 3 y 4).
 |
| |
FIGURA
1. Actividad sanguínea de GSH-Px (x ± DE ) en vacas selenio-deficientes
(Se-D) y seleniosuplementadas (Se-S) con selenato de bario (día 0). Ambos
grupos mantenidos con ración selenio-deficiente. |
-
Período de partos entre 45 y 60 días postsuplementación.
* P < 0.05 entre grupos.
Blood GSH-Px activity (mean ± SD) in cows
fed a selenium deficient diet (Se-D) and cows supplemented (Se-S) with
barium selenate on day 0. |
CUADRO
1. Concentraciones séricas de IgG, IgM e IgA (X ± DE en g/l), 6 y 2 semanas
antes del parto, en vacas selenio-deficientes (Se-D) y selenio-suplementadas
(Se-S). |
Serum IgG, IgM and
IgA concentrations (mean ± SD in g/l) on weeks 6 and 2 before partum in
selenium deficient (Se- D) and selenium supplemented (Se-S) cows. |
| |
| |
IgG total |
IgM
|
IgA |
Semana |
6 |
2 |
6 |
2 |
6 |
2 |
Grupo
Se-S
Se-D |
51.2 ± 5.4a
43.7 ± 7.0a
|
27.2 ± 9.0b
32.3 ± 7.0b
|
3.09 ± 0.9a
3.41 ± 1.6a
|
2.94 ± 0.79a
3.34 ± 1.52a
|
0.19 ± 0.12a
0.16 ± 0.08a
|
0.17 ± 0.09a
0.12 ± 0.05a
|
| |
| a Letras distintas en las filas señalan diferencias
significativas entre el período (P < 0.05). |
CUADRO
2. Concentraciones
de IgG, IgG1, IgM e IgA (X ± DE en g/l) en el calostro de vacas selenio-deficientes
(Se-D) y selenio-suplementadas (Se-S). |
IgG, IgG1, IgM e IgA
colostrum concentrations (mean ± SD in g/l) in selenium deficient (Se-D)
and selenium supplemented (Se-S) cows. |
| |
| Grupo |
IgG Total |
IgG1 |
IgM |
IgA |
Se-S
Se-D |
124.6 ± 35.5
123.2 ± 43.1 |
84.4 ± 12.3a
103.0 ± 12.5b |
4.97 ± 0.97
6.05 ± 1.72 |
5.81 ± 1.68
4.58 ± 1.57 |
| |
a Letras distintas
en las columnas señalan diferencias significativas entre grupos
(P < 0.05). |
CUADRO
3. Concentraciones séricas (X ± DE en g/l) de IgG e IgG1 el día del nacimiento
y a los 2, 4 y 6 días de edad en terneros nacidos de las vacas selenio-deficientes
(Se-D) y selenio-suplementados (Se-S). |
IgG and IgG1 serum
concentrations (mean ± SD in g/l) at the day of birth, 2, 4 and 6 days
of age in calves of selenium deficient (Se-D) and selenium supplemented
(Se-S) cows. |
| |
| - |
IgG Total |
IgG1 |
| Edad (días) |
0 |
2 |
4 |
6 |
0 |
2 |
4 |
6 |
Grupo
Se-S
Se-D |
1.86 ± 2.02a
1.78 ± 0.54a
|
28.6 ± 12.6b
26.6 ± 20.0b
|
29.9 ± 12.9b
31.7 ± 18.2b
|
28.6 ± 12.4b
34.5 ± 16.9b
|
1.63 ± 2.35a
0.87 ± 0.36a
|
17.7 ± 6.54b
16.1 ± 10.6b
|
17.8 ± 6.54b
17.6 ± 9.02b
|
17.5 ± 7.11b
17.0 ± 9.32b
|
| |
a Letras distintas en
la fila de cada grupo señalan diferencias significativas entre días
postnacimiento (P < 0.05).
Edad 0: Concentración determinada en la primera hora desde el nacimiento. |
CUADRO
4. Concentraciones séricas (X ± SD en mg/dl) de IgM e IgA el día del nacimiento
y a los 2, 4 y 6 días de edad en terneros nacidos de las vacas selenio-deficientes
(Se-D) y selenio-suplementados (Se-S). |
IgM and IgA serum concentrations
(mean ± SD in mg/dl) at the day of birth, 2, 4 and 6 days of age in calves
of selenium deficient (Se-D) and selenium supplemented (Se-S) cows. |
| |
| - |
IgM |
IgA |
| Edad (días) |
0 |
2 |
4 |
6 |
0 |
2 |
4 |
6 |
Grupo
Se-S
Se-D |
20 ± 16 a
37 ± 3 a
|
|
144 ± 62 b
163 ± 75 b |
103 ± 34 b
142 ± 63 b
|
7 ± 0 a
7 ± 0 a
|
125 ± 58 b
78 ± 40 b
|
87 ± 43 b
60 ± 20 b
|
63 ± 36 b
48 ± 12 b
|
| |
a Letras distintas en la fila de cada grupo señalan
diferencias significativas entre días postnacimiento (P < 0.05).
Edad 0: Concentración determinada en la primera hora desde el nacimiento. |
DISCUSION
Los requerimientos de Se para
bovinos corresponden a 0.1 ppm/Se (base materia seca) y de 0.3 ppm/Se/MS en
el caso de vacas en lactancia (NRC, 2001). La concentración mínima que debe
tener la dieta de bovinos es de 0.05 ppm/ Se/MS, ya que dietas con concentraciones
menores a esta cifra se asocian con alteraciones en la salud y producción del
ganado (Underwood and Suttle, 1999). La dieta utilizada en
este experimento contenía 0.04 ppm/Se/MS, lo que permitió predisponer a la presentación
de problemas asociados a la deficiencia de Se en los animales no suplementados.
No obstante, no se observaron alteraciones atribuibles a esta deficiencia durante
el período de partos y el puerperio.
La actividad de la enzima GSH-Px
se considera un buen indicador del estatus de selenio en rumiantes (Stowe
y Herdt, 1992) y el selenato de bario una fuente adecuada de aporte de selenio.
Además, este producto tiene la ventaja que mantiene los niveles de Se durante
varios meses debido a su lenta liberación desde el sitio de inyección (Lee
y col., 1999). En este estudio, la suplementación de las vacas con selenato
de bario aumentó significativamente la concentración de GSH-Px a partir del
día 30 postsuplementación, alcanzando niveles superiores a 200 UI/ g/Hb durante
el período de los partos, apropiados para
vacas en lactancia (Ceballos y Wittwer, 1996). Contrario a
ello, la actividad de GSH-Px de las vacas del grupo Se-D se mantuvo dentro de
lo considerado bajo marginal o deficientes (< 100 U/g Hb), corroborando con
ello la deficiencia de Se en la dieta. Este hecho permitió establecer las diferencias
y similitudes entre grupos.
La disminución de la concentración
de IgG total en las vacas de ambos grupos durante el fin de la gestación coincide
con el transporte activo de inmunoglobulina IgG desde la circulación sanguínea
a la glándula mamaria (Larson y col., 1980). Llama la atención
que, aun cuando la concentración de IgG total en calostro de las vacas no presentó
diferencias entre los grupos suplementados y no suplementados, los resultados
muestran que la suplementación con Se afecta negativamente la concentración
de IgG1 en el calostro. Si se considera que toda la IgG, la mayor parte de la
IgM y cerca del 50% de la IgA del calostro bovino provienen del plasma (Tizar,
1998), estos resultados sugieren que el Se favorece el aumento de la concentración
de IgG2 en la glándula mamaria. Esto podría explicar por qué no se observaron
diferencias significativas en la concentración total de IgG entre los grupos
experimentales. El aumento de IgG2 inducido por el Se sería de gran importancia,
ya que los neutrófilos de bovino presentan receptores solamente para IgG2 (Howard
y col., 1980), lo cual favorece la defensa contra las infecciones bacterianas.
Este aspecto requiere mayor estudio.
La ausencia de un efecto favorable
de la suplementación con Se sobre la inmunidad pasiva de los terneros del grupo
Se-S concuerda con estudios previos realizados por Lacetera y
col. (1996), quienes observaron que la suplementación de Se asociado a la
vitamina E favorecen la producción de calostro y leche de las vacas, pero no
benefician la inmunidad pasiva de las crías. También se ha señalado que en vacas
de aptitud cárnica, a mitad de preñez y mantenidas en praderas selenio-deficientes,
la administración parenteral de Se asociado a vitamina E no afecta el contenido
de IgG en calostro (Swecker y col., 1995); sin embargo, estos
mismos autores señalan que el uso de Se solo o asociado con vitamina E, pero
administrado con una mezcla mineral, aumenta las concentraciones de IgG en el
calostro de las vacas y en el plasma sanguíneo de sus crías.
En nuestro estudio no se observaron
diferencias para IgM e IgA entre grupos. Sin embargo, otras investigaciones
realizadas in vitro han demostrado un efecto positivo del
Se sobre plasmocitos de la glándula mamaria encargados de la producción local
de inmunoglobulinas (Wuryastuti y col., 1993; Hogan
y col., 1993).
Las inmunoglobulinas detectadas
en los terneros corresponden a las absorbidas desde el calostro, ya que la IgM,
IgA, IgG2 e IgG1 producidas por el neonato aparecen sólo a los 4, 4, 8 y 32
días de vida, respectivamente (Husband y Lascelles, 1975).
La no observación de diferencias en las concentraciones séricas de inmunoglobulinas
entre los terneros de ambos grupos de vacas en los primeros seis días de vida
son consecuentes con la ausencia de diferencias entre las concentraciones de
inmunoglobulinas en el suero y calostro de los grupos Se-S y Se-D. Esta observación
sugiere que el balance positivo de Se en las vacas suplementadas no jugaría
un rol importante en la absorción de inmunoglobulinas por la mucosa intestinal
del ternero.
La ausencia de diferencias significativas
entre ambos grupos experimentales, en este estudio, así como, los resultados
de estudios previos sobre inmunidad y Se revisados por Finch y
Turner (1996), demuestran que la respuesta de anticuerpos tanto en animales
selenio-deficientes como en suplementados con selenio es variable y en algunos
casos hasta contradictoria. Esto sugiere que la modulación de la respuesta inmune
por este microelemento estaría influenciada por diversos factores, tales como
la vía de administración (Swecker y col., 1995) y las interacciones
entre nutrientes, como se ha señalado para vitamina E y Se (Kubena
y McMurray, 1996). Estos aspectos habitualmente no son considerados cuando
se indica la suplementación con Se. En este sentido, es necesario realizar estudios
tendientes a establecer protocolos de utilización de Se de acuerdo a objetivos
bien definidos.
Los resultados obtenidos en el
presente estudio permiten concluir, en primer lugar, que la deficiencia nutricional
de Se en vacas gestantes no afecta la concentración de inmunoglobulinas totales
en calostro ni en el plasma sanguíneo de las crías; en segundo lugar, el uso
de selenato de bario en vacas gestantes selenio-deficientes restablece el balance
nutricional de Se; sin embargo, disminuye la concentración de IgG1 sin afectar
los niveles de IgG total en el calostro. Este hecho implicaría un aumento de
IgG2, lo cual tiene gran importancia, ya que los neutrófilos de rumiantes sólo
poseen receptores para IgG2. Esta condición favorece la función de los neutrófilos
y tiene especial relevancia en el control de las infecciones bacterianas.
BIBLIOGRAFIA
CEBALLOS, A.,
F.G. WITTWER. 1996. Metabolismo del selenio en rumiantes. Arch. Med.
Vet. 28: 5-18.
CEBALLOS, A.,
F. WITTWER, P. A CONTRERAS, H. BÖHMWALD. 1998. Actividad sanguínea de glutatión
peroxidasa en rebaños lecheros a pastoreo: variación según edad y época del
año. Arch. Med. Vet. 30: 13-22.
CEBALLOS, A.,
F. WITTWER, P.A. CONTRERAS, E. QUIROZ, H . BÖHMWALD. 1999. Actividad de glutation
peroxidasa en bovinos a pastoreo correlacionada con la concentración sanguínea
y plasmática de selenio. Pesq. Agrop. 34: 2331-2338.
CONTRERAS, P.A.,
R. MATAMOROS, R. MONROY, J. KRUZE, V. LEYAN, M. ANDAUR, H. BÖHMWALD, F. WITTWER.
2002. Effect of a selenium deficient diet on blood values of T3 and
T4 in cows. Comp. Clin. Path. 11: 65-70.
EDMONSON, A.J.,
I.J. LEAN, L.D. WEAVER, T. FARVER, G. WEBSTER. 1989. A body condition scoring
chart for Holstein dairy cows. J. Dairy Sci. 72:
68-78.
FINCH, J..M.,
R. J. TURNER. 1996. Effects of selenium and vitamin E on the immune responses
of domestic animals. Res. Vet. Sci. 60: 97-106.
HOGAN, J.S.,
W.P. WEISS, K.L SMITH. 1993. Role of vitamine E and selenium in host defense
against mastitis J. Dairy Sci. 76: 2795-2803.
HOWARD, C.J.,
TAYLOR, G., J. BROWNLIE. 1980. Surface receptors for immunoglobulin on bovine
polymorphonuclear neutrophils and macrophages. Res. Vet. Sci.
29: 128-130.
HUSBAND, A.J.,
A.K. LASCELLES. 1975. Antibody response to neonatal immunization in calves.
Res. Vet. Sci. 18: 201-207.
KRUZE, J.,
A. MELLA, P. A. CONTRERAS, F. WITTWER, V. LEYAN, R. MATAMOROS.2000. Experimental
Staph. aureus mastitis in selenium-deficient and selenium-supplemented
dairy cows. XXI Congreso Mundial de Buiatría. Punta del Este-Uruguay.
KUBENA, K.S.,
D.N. Mc MURRAY. 1996. Nutrition and the immune system: a review of nutrientnutrient
interactions. J. Am. Diet. Assoc. 96: 1156-1164.
LACETERA,
N., U. BERNABUCCI, B. RONCHI, A. NARDONE. 1996. Effects of selenium and vitamin
E administration during a late stage of pregnancy on colostrum and milk production
in dairy cows, and on passive immunity and growth of their offspring. Am.
J. Vet. Res. 57: 1776-1780.
LARSON, B.L.,
J.L HEARY, J.E. DEVERY. 1980. Immunoglobulin production and transport by the
mamary gland J. Dairy Sci. 63: 665-671.
LEE, J., D.G
MASTERS, C.L WHITE, N.D. GRACE, G.J. JUDSON. 1999. Current issues in trace element
nutrition of grazing livestock in Australia and New Zealand. Aust.
J. Agric. Res. 50: 1341-1364.
LOPEZ ALONSO,
M., M. MIRANDA, J. HERNANDEZ, C. CASTILLO, J. L. BENEDITO. 1997. Glutatión peroxidasa
en las patologías asociadas a deficiencias de selenio en rumiantes. Arch.
Med. Vet. 29: 171-179.
MATAMOROS,
R., P. A. CONTRERAS, F. WITTWER, I.M. MAYORGA. 2003. Hipotiroidismo en rumiantes.
Arch. Med. Vet. 35: 1-11.
NATIONAL RESEARCH
COUNCIL, NRC. 2001. Nutrient requirements of dairy cattle. 7th Ed. National
Academy Press, Washington, DC.
PESUTIC, D.,
V. LEYAN, G. SCHURIG, F. WITTWER, P. CONTRERAS, J. KRUZE, R. MATAMOROS. 2001.
Efectos de una dieta selenio-deficiente sobre la respuesta inmune a la vacuna
RB51 en vaquillas a pastoreo. V Jornadas Chilenas de Buiatría. Puerto Varas-Chile.
pp. 141-142.
STOWE, H.
D., T.H. HERDT. 1992. Clinical assessment of selenium status of livestock. J.
Anim. Sci. 70: 3928-3933
SWECKER, W.,
C. THATCHER, D. EVERSOLE, D. BLODGETT, D. SCHURIG. 1995. Effect of selenium
supplementation on calostral IgG concentration in cows grazing selenium-deficient
pastures and on postsuckle serum IgG concentration in their calves. Am.
J. Vet. Res. 56: 450-453.
TIZAR, I.
R. 1998. Inmunología Veterinaria. McGraw Hill Interamericana editores. México.
UNDERWOOD,
E.J., N F. SUTTLE. 1999 The Mineral Nutrition of Livestock. 3rd Ed. Pp. 343-373.
CAB International, Wallingford, Oxford, UK.
WITTWER, F.,
O. ARAYA, P.A. CONTRERAS. 2002. Strategic procedures for important herd problems
of heifers. In: Kaske M., Scholz M, Höltershinken M (eds.) Recent Developments
and Perspectives in Bovine Medicine. Pp. 396-409 Keynote Lectures, XXII World
Buiatric Congress. Hannover, Germany.
WURYASTUTI,
H., H.D. STOWE, R.W. BULL, E.R. MILLER. 1993. Effects of vitamin E and selenium
on immune response of peripheral bood, colostrum and milk leukocytes of sows.
J. Anim. Sci. 71: 2464-2472.
ZAR, J.H. 1996.
Biostatistical Analysis. 3rd Ed. Prentice Hall. New Jersey. USA.
Aceptado: 06.07.2004.