Bosque, Vol. 24 N° 1, 2003, pp. 35-45
ARTICULOS
Utilización del modelo 3-PG, un modelo basado en procesos, en el análisis de la productividad de plantaciones de pino radiata manejado con diferentes regímenes silviculturales
Using the 3-PG in the analysis of productivity of radiata pine plantations under different silvicultural regimes
ROLANDO RODRIGUEZ LEIVA 1, MIGUEL ESPINOSA BANCALARI 2, PEDRO REAL HERMOSILLA 3
1
CONAF, Barros Arana 215, 2º piso, Concepción, Chile.
2 Universidad de Concepción, Facultad de Ciencias
Forestales, Departamento de Silvicultura, Casilla 160-C, Concepción, Chile.
3 Universidad de Concepción, Facultad de Ciencias
Forestales, Departamento de Manejo de Recursos Forestales, Casilla 160-C,
Concepción, Chile.
Summary
The effects of both crown structure and fertilization regime on productivity were studied. We compared the growth and development of stands managed with silvopastoral methods with those of a traditionally managed stand. A process-based model, designated 3-PG and validated by stem analysis, was used to estimate growth. We concluded that the leaf area index was strongly controlled by the design of the plantation and its final density. The fertilization regime had a positive effect on the leaf area index but its potential was limited by crown structure. Under the traditional forestry regime, the increase in stem-wood biomass was strongly related to the leaf area index. However, in stands under silvopastoral management, the increase is attributable to changes in allocation from the biomass of the fine roots, as an effect of the fertilization regime. Because of the effect of thinning on crown structure, the final wood production was more affected by thinning than by the availability of the site resources. Therefore, maximization of wood production will depend upon the combined management of site resources with the intensity of and opportunity for thinning.
Key words: simulation, site occupancy, productivity, radiata pine, 3-PG model.
Resumen
Se estudia el efecto de la estructura de copas y el régimen de fertilización en la producción de madera. Para ello se comparó el crecimiento y desarrollo de dos rodales, uno manejado con un sistema silvopastoral con respecto a uno manejado en forma tradicional para la producción de madera. Para estimar el crecimiento se utilizó un modelo basado en procesos denominado 3-PG, que se validó con un análisis fustal. Se concluyó que el índice de área foliar fue controlado fuertemente por el diseño de plantación y la densidad final. El régimen de fertilización tuvo efectos positivos sobre el índice de área foliar, pero su potencial fue limitado por la estructura de copas. En el régimen forestal el incremento en biomasa fustal se presentó estrechamente relacionado con el índice de área foliar. Sin embargo, en rodales con manejo silvopastoral, su incremento se atribuye a un cambio de localización desde la biomasa de raíces finas, por efecto del régimen de fertilización. El raleo, por su efecto en la estructura de copas más que la disponibilidad de los recursos del sitio, afecta la producción final de madera. Por tanto, la maximización de la producción de madera dependerá del manejo conjunto de los recursos del sitio con la intensidad y oportunidad del raleo.
Palabras claves: simulación, ocupación del sitio, productividad, pino radiata, Modelo 3-PG.
INTRODUCCION
La producción de madera en cultivos forestales depende de la tasa de asimilación neta y de la localización relativa de los fotosintatos asignados al crecimiento fustal (McMurtrie et al. 1990). Además de los elementos del clima como radiación solar y régimen de temperaturas, la fotosíntesis dependerá de la producción de follaje y de la tasa de asimilación de carbono por unidad de área o masa foliar (Wang y Jarvis 1990); la ganancia de carbono está determinada por la disponibilidad de recursos del sitio, tales como luz, agua y nutrientes (Carlyle 1995). Sin embargo, debido a la relación inversa entre la eficiencia de crecimiento y el índice de área foliar, el tamaño y estructura de las copas y la densidad del rodal influencian tanto la ganancia como la distribución del carbono (Perry 1985, Roberts et al. 1993). Por consiguiente, la producción de madera es función de la estructura del rodal, la disponibilidad de recursos del sitio y limitantes climáticas (Waring y Runing 1998). Al respecto, Perry (1985) y Cannel (1989) han señalado que la masa foliar y la estructura de copas, más que los recursos del sitio, determinan la producción de madera. A su vez, McMurtrie y Landsberg (1992) han determinado para pino radiata que la disponibilidad de recursos como agua y nutrientes parecen ser más importantes en la producción de fotosintatos y su localización en el fuste que la estructura del rodal.
Para el análisis de los procesos que afectan la ganancia en carbono y su localización en los distintos componentes del árbol y su potencial utilidad como herramienta de investigación en el manejo silvícola, han surgido los modelos de simulación basados en procesos, que pueden convertirse en un valioso instrumento de apoyo en la toma de decisiones en el contexto del desarrollo forestal sustentable (Mäkelä et al. 2000a). Estos modelos se basan en la parametrización tanto del ambiente físico como de los procesos fisiológicos (Landsberg y Hingston 1996, Waring y Runing 1998) y en la interdependencia de los ciclos del agua, carbono y nutrientes (Runing y Gower 1991, McMurtrie y Landsberg 1992).
De los diversos modelos existentes basados en procesos, uno de los más utilizados es el 3-PG, desarrollado por Landsberg y Waring (1997). Este modelo está siendo aplicado en Estados Unidos, principalmente en la utilización forestal de bosques naturales, y en el análisis del efecto ambiental de las plantaciones en la producción de agua en importantes cuencas hidrográficas en Sudáfrica (Mäkelä et al. 2000a).
En este estudio se utilizó el modelo 3-PG en el análisis de la productividad de rodales de pino radiata, creciendo bajo dos regímenes silvícolas. En particular, se plantea la hipótesis de que el diseño de plantación y la densidad determinada por el régimen de raleos, al afectar el índice de área foliar potencial máximo, se constituye por sobre el manejo de los recursos del sitio en el factor más importante que explica la producción de madera.
MATERIAL Y METODOS
Area de estudio y diseño del experimento. Un experimento silvopastoral fue instalado por la Corporación Nacional Forestal, en la localidad de Tanumé, comuna de Pichilemu (VI Región), Chile (34º9-34º15 S; 72º53-72º59 W). Los suelos del área se clasifican dentro del orden Alfisol, suborden Xeralfs (Rodríguez et al. 2001). El clima es templado subhúmedo, con cuatro a seis meses de sequía y una nubosidad abundante por su cercanía al mar. El área de estudio, para el período 1983- 1999, presenta una precipitación media anual de 702,7 mm, con una temperatura media mínima de 8,6ºC y temperatura media máxima de 15,4ºC, de acuerdo con datos obtenidos de la estación meteorológica de Tanumé, distante 2,5 km de la zona de estudio.
El experimento silvopastoral se estableció a inicios de 1983 y consistió en una plantación de pino radiata con una densidad inicial de 1.000 árboles ha1 en un rodal de 6 ha, la que se diseñó mediante un arreglo en franjas con distanciamiento entre árboles de 2 x 3 m y de 7 m entre franjas. En 1985 se estableció artificialmente la pradera entre las franjas con pastos, la que fue fertilizada anualmente y el rodal pasó a constituir un módulo silvopastoral. En un área contigua se estableció una plantación con fines de producción de madera a una densidad inicial de 1.600 árb ha1 y espaciamiento de 2,5 x 2,5 m (cuadro 1). Las intervenciones silvícolas, en ambos rodales, correspondieron a raleos los años 1989, 1991 y 1993 y a podas los años 1988, 1990 y 1993. El criterio utilizado para definir la oportunidad, intensidad y densidades residuales se orientó a la optimización del sistema silvopastoral, tratando de buscar un equilibrio que permitiera el aprovechamiento conjunto de los árboles y de la pradera.
Modelo 3-PG. El modelo 3-PG es un modelo de crecimiento basado en procesos (figura 1). De acuerdo a la descripción de Landsberg y Waring (1997), el modelo tiene la siguiente estructura:
Asignación de
tratamientos de acuerdo con el régimen de manejo, densidad inicial,
diseño de plantación y tipo de pradera en pino radiata de 16 años.
Treatment assignment according to management regime, initial density, and prairie type, for 16-year-old radiata pine. |
Régimen
de manejo |
Símbolo |
Densidad (árboles ha-1) |
Diseño
de plantación |
Tipo
de pradera |
Silvopastoral Forestal |
T1 T2 |
1.000 (185) 1.600 (489) |
Hileras Normal |
Artificial Natural |
Nota: El número en paréntesis indica la densidad actual. |
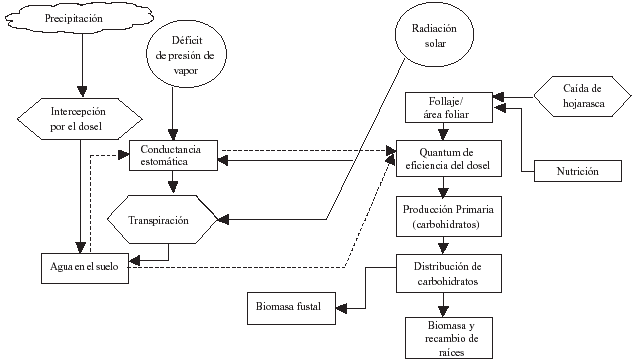
Figura
1: Esquema simplificado del modelo 3-PG para predecir el
crecimiento de un bosque. (Coops et al. 1998).
Schematic representation of a simplified 3-PG model to predict forest growth. |
i) Estima la Producción
Primaria Bruta (PG) a partir de la radiación fotosintéticamente
activa utilizable (![]() p.a.u)
y el quantum de eficiencia del dosel (
p.a.u)
y el quantum de eficiencia del dosel (![]() C).
El valor de
C).
El valor de ![]() p.a.u
se obtiene por una reducción del valor de la radiación fotosintéticamente
activa (
p.a.u
se obtiene por una reducción del valor de la radiación fotosintéticamente
activa (![]() p.a.)
a través de factores modificadores adimensionales cuyos valores varían
entre 0 y 1. Los coeficientes modificadores reflejan las condiciones impuestas
sobre
p.a.)
a través de factores modificadores adimensionales cuyos valores varían
entre 0 y 1. Los coeficientes modificadores reflejan las condiciones impuestas
sobre ![]() p.a.u
debido al cierre de estomas, causados por altos niveles de déficit de
presión de vapor (D), sequías hídricas y estado nutricional del suelo.
La sequía del suelo está definida por la proporción de agua en la zona
radicular con respecto a la máxima cantidad de agua disponible (
p.a.u
debido al cierre de estomas, causados por altos niveles de déficit de
presión de vapor (D), sequías hídricas y estado nutricional del suelo.
La sequía del suelo está definida por la proporción de agua en la zona
radicular con respecto a la máxima cantidad de agua disponible (![]() )
o por temperaturas de congelamiento.
)
o por temperaturas de congelamiento.
ii) Estima la Productividad Primaria Neta (PN) a partir de PG. Para ello el modelo utiliza la proporción PN/PG=Cpp, la que se ha estimado en 0,47 ± 0,04 SD. Este valor ha demostrado ser relativamente constante para diversos tipos de bosques y localidades geográficas (Waring et al. 1998).
iii) Para estimar el carbono localizado en la biomasa de raíces, utiliza relaciones simples derivadas de información proporcionada por la literatura actual para evaluar el crecimiento radicular, la tasa de recambio de raíces finas y los efectos de las condiciones del sitio sobre el crecimiento radicular. Para ello se han considerado dos relaciones básicas: i) la relación inversa entre el crecimiento fustal y la fracción de PN localizada en el suelo (Beets y Whitehead 1996) y, ii) el efecto ambiental causado por condiciones de sequías, régimen de temperatura y nutrición, que incrementan la localización anual del carbono en biomasa de raíces entre un 25% y 60% (Santantonio 1989, Beets y Whitehead 1996).
iv) El modelo 3-PG utiliza un submodelo, que ha sido derivado de la ley de autorraleo y de las tasas de crecimiento fustal, que permite calcular los cambios en la densidad del rodal con el tiempo (ley de 3/2 o de autorraleo).
v) Para determinar la localización del carbono en los diversos componentes del árbol como follaje, fuste y raíces, se utilizan relaciones alométricas, de la forma:
Wi = aiWNi
En que W es la biomasa total del árbol e i es el componente del árbol. El parámetro Ni refleja las características genéticas de la especie.
vi) Para estimar el efecto del decrecimiento de la biomasa aérea del bosque con la edad, el modelo usa la relación lineal entre conductividad hidráulica y PN (Mencuccini y Grace 1996). Esta relación sustenta la hipótesis de que la conductancia del fuste declina con la edad induciendo una menor gc. Para incorporar este efecto, Landsberg y Waring (1997) utilizaron una función de decrecimiento de la biomasa aérea con la edad.
Antecedentes del sitio para la aplicación del Modelo 3-PG. Las variables temperatura máxima, mínima y media mensual, entre los años 1983 y 1999, se obtuvieron de la estación meteorológica ubicada en el predio Tanumé, contigua al área de estudio. El déficit de presión de vapor mensual se estimó mediante la diferencia de presión de vapor saturado (Pvs), utilizando la temperatura máxima (T1) y mínima mensual (T2) en ºC, de acuerdo a la fórmula de Runing et al. (1987), en que:
Pvs, kPa = 0,61078 * exp(17,269*(T1/T2+273,3) (1)
La radiación solar diaria se derivó de los datos de temperatura máxima y mínima de Tanumé, de acuerdo al procedimiento descrito por Bristow y Campbell (1984). Estos valores fueron corregidos con antecedentes de localización, latitud, longitud y altitud y considerando una superficie plana del área de estudio, obteniéndose el máximo potencial de radiación solar. Este potencial se corrigió mediante la variación mensual del ángulo de declinación solar sobre la superficie terrestre (Waring 2000). La influencia de nubes y la constante neblina en el área de estudio, por su cercanía al mar, reduce el potencial de radiación solar. Para estimar esta reducción se utilizaron las ecuaciones desarrolladas por Bristow y Campbell (1984) y corregidas por Waring (2000). Se asumió un valor 0,15 para la proporción entre la radiación difusa y directa, valor que es considerado constante durante el año. Los valores mensuales de las principales variables meteorológicas para el sitio se entregan en la figura 2.
En el empleo del modelo
se asumió que la radiación solar extraterrestre es el doble de ![]() p.a
y que la fracción de radiación absorbida por el dosel es función
del índice de área foliar (L), el que se calcula mediante las leyes de
Beer (Landsberg y Waring 1997). La respuesta de la conductancia estomática
(gc) a la humedad ambiental se calcula en relación con el déficit
de presión de vapor (cuadro 2).
p.a
y que la fracción de radiación absorbida por el dosel es función
del índice de área foliar (L), el que se calcula mediante las leyes de
Beer (Landsberg y Waring 1997). La respuesta de la conductancia estomática
(gc) a la humedad ambiental se calcula en relación con el déficit
de presión de vapor (cuadro 2).
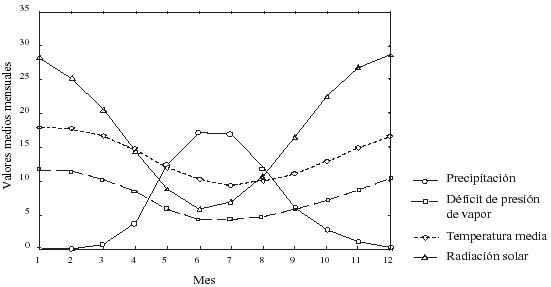
Figura
2: Valores medios mensuales de Precipitación (cm), Déficit
de presión de vapor (mb), Temperatura media (ºC) y Radiación solar
(MJm2), para el sitio Tanumé en el período 1983-1999.
Total monthly precipitation (cm), mean daytime atmospheric vapor pressure deficit (mb), mean monthly temperature (ºC), total daily solar radiation (MJ/m2), for the Tanumé site, in the period 19831999. |
Modelos de las
principales funciones y parámetros utilizados en este estudio, los
cuales pueden diferir de la versión original del modelo 3-PG (Landsberg
y Waring, 1997), de acuerdo con las modificaciones de Sands (2000),
en pino radiata de 16 años de edad. Model functions and parameters used in this study of 16-year-old radiata pine, which differ in several ways from the original version of 3-PG (Landsberg and Waring 1997), according to Sands (2000). |

| Nota:
L es el índice de área foliar y se define como la superficie de
acículas por unidad de superficie del suelo, m2 m2;
|
Los efectos de la temperatura
de congelamiento y del régimen de fertilización en la fotosíntesis fueron
modificados en su versión original por Sands (2000). Los autores originales
del modelo asumieron que para un día cualquiera, si la temperatura cae
bajo 0ºC no hay fotosíntesis (Landsberg y Waring 1997). Además, se considera
que en la medida que la proporción ![]() p.a.u/
p.a.u/![]() p.a.u
tiende a 1, el suelo ofrece las mejores condiciones nutricionales para
el crecimiento, en ausencia de otros factores limitantes. Por ello, esta
relación se modifica mediante una función que incorpora el efecto del
régimen de fertilización (fN(FR)). El modificador
es un ranking que varía entre FR=0 para suelos sin fertilización y FR=1,
en suelos fertilizados (Sands 2000).
p.a.u
tiende a 1, el suelo ofrece las mejores condiciones nutricionales para
el crecimiento, en ausencia de otros factores limitantes. Por ello, esta
relación se modifica mediante una función que incorpora el efecto del
régimen de fertilización (fN(FR)). El modificador
es un ranking que varía entre FR=0 para suelos sin fertilización y FR=1,
en suelos fertilizados (Sands 2000).
Las modificaciones a las funciones para la caída de hojarasca, la localización a biomasa de raíces y fuste, son explicadas por Sands (2000). Las funciones y parámetros utilizados se entregan en el cuadro 2. La cantidad de agua disponible se estimó a partir de curvas de retención de agua en el perfil del suelo. Puesto que la ley de autorraleo forma parte del modelo, no es factible simular directamente el efecto del raleo en el desarrollo del rodal. Por ello, las simulaciones se realizaron considerando las densidades evaluadas directamente antes y después del último raleo, en cada rodal.
ESTIMACION DEL CRECIMIENTO MEDIANTE ANALISIS FUSTAL
Muestreo. El trabajo de terreno se desarrolló entre diciembre de 1999 y abril de 2000. En parcelas permanentes instaladas por la Corporación Nacional Forestal, en ambos rodales, se midieron diámetros a la altura del pecho (Dap) de todos los árboles para obtener la amplitud diamétrica. Luego se seleccionaron mediante muestreo aleatorio simple 9 árboles en cada rodal y 18 árboles en total. Previo al volteo de cada individuo se midió el Dap. En el árbol volteado, su altura total (H), incluyendo la altura del tocón. Posteriormente, se cortaron rodelas de 2 a 3 cm de espesor a la altura del tocón (0,30 m) del Dap y sobre éste a distancias de 3,5 m incluyendo además una rodela en la base de la copa viva. Las rodelas fueron etiquetadas, guardadas en bolsas plásticas y almacenadas en cámara de frío a 2ºC para su análisis posterior.
Análisis fustal. En laboratorio, las rodelas después de saturarse en agua fueron fotografiadas, para lo cual cada una de ellas se ubicó sobre una plataforma de aluminio con un pedestal para fijar una cámara digital Epson, modelo Photo PC 750Z. Las mediciones se realizaron sobre fotografías digitales, las que fueron georreferenciadas mediante el software Arcinfo NT 7,2. Mediante la ayuda de referencias fotografiadas en conjunto con la rodela en análisis, se logró determinar la escala de la imagen y su posterior procesamiento. Con el software Adobe Photoshop 4,0 para procesamiento de imágenes se superpuso cada imagen digital de la rodela a un sistema de referencia de coordenadas x e y, en que el origen se ubicó en la médula. Por tanto, las rodelas quedaron divididas en cuatro cuadrantes, lo que permitió medir en forma independiente la distancia entre anillos con el software Arc View 3.0. Estas distancias se corrigieron por un factor de escala que corregía el efecto del grosor de la rodela. Con los resultados de estas mediciones se obtuvieron los diámetros a distintas alturas del fuste de cada uno de los árboles muestreados para los períodos anuales de crecimiento comprendidos entre 1990 y 1999. El volumen total del árbol, en cada período de crecimiento, se calculó de acuerdo a la forma geométrica de la porción del fuste: el volumen del tocón como un cilindro, la sección a la altura del Dap como el tronco del niloide, las secciones entre el Dap y la base de la copa viva como tronco de parábola y de la base de la copa viva al ápice como un paraboloide (Husch et al. 1982). Para la estimación del volumen del rodal se utilizó un modelo de razón en base al área basal (Van Laar y Akça 1997), para lo cual se emplearon las variables de estado del rodal, densidad, Dap y área basal, las que fueron medidas anualmente en 22 parcelas permanentes (11 parcelas por tratamiento) entre 1990 y 1999. Para determinar la densidad de la madera las rodelas se sumergieron en agua para obtener su volumen fresco y posteriormente fueron secadas a 100ºC por 48 horas o hasta obtener un peso constante. Los estadísticos utilizados para la validación del modelo 3-PG fueron como medida de precisión la raíz del error cuadrático medio (REMC) y su expresión porcentual y como medida del sesgo la Diferencia Agregada (DIFA) en unidades de medición y porcentaje.
RESULTADOS Y DISCUSION
Validación del modelo en la predicción del crecimiento en volumen. La primera fase de la simulación fue evaluar la capacidad del modelo para predecir el crecimiento en volumen. Las curvas de crecimiento en volumen, estimado mediante análisis fustal y los simulados por el 3-PG, en ambos tratamientos, se presentan en la figura 3. Las estimaciones más precisas el modelo las alcanzó en T2 presentando ambos tratamientos sesgos mínimos (cuadro 3).
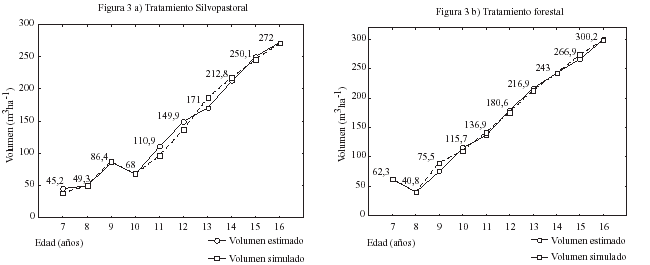
Figura
3: Diagrama de dispersión de los volúmenes observados y
simulados (m3 ha1) por el modelo 3-PG para
los distintos tratamientos en pino radiata de 16 años. Dispersion diagram of the observed and simulated volumes (m3 per ha) calculated by the 3-PG model for different treatments, in 16-year-old radiata pine. |
Medidas de precisión
y sesgo para la validación del modelo 3-PG en pino radiata de 16
años de edad. Precision and bias measurements for validation of the 3-PG model in 16-year-old radiata pine. |
Tratamiento |
Medidas
de precisión |
Medidas
de sesgo |
||
REMC Valor Absoluto |
(%) |
DIFA Valor Absoluto |
(%) |
|
T1 |
3,3 |
4,4 |
0,8 |
0,5 |
T2 |
5,5 |
2,9 |
0,7 |
0,4 |
Nota:
REMC es la raíz del error medio cuadrático en valores absolutos
y en porcentaje. DIFA es la diferencia agregada en unidades de medición
y porcentaje. |
Efecto del régimen de manejo y el raleo en el desarrollo del índice de área foliar y la productividad. Los resultados obtenidos mediante simulación para el desarrollo de L en los distintos tratamientos se presentan en la figura 4a.
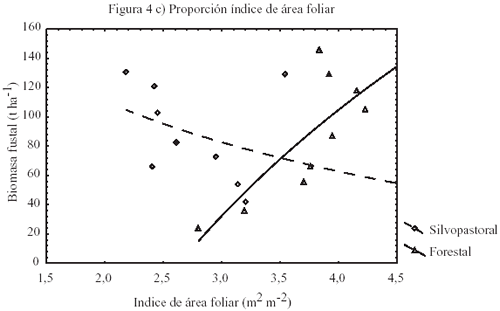
El desarrollo de L en T2 sigue el comportamiento típico del efecto del raleo en plantaciones. Esto es, un incremento desde valores bajos previo al raleo para alcanzar en el tiempo un máximo y luego decrecer con el cierre de copas. En este rodal, L alcanzó un máximo de 4,2 m2 m2 (cuadro 4) después de 4 años. En cambio, en el rodal con manejo silvopastoral el máximo L fue de 3,5 m2 m2 (cuadro 4) y se alcanzó a los 2 años para luego decrecer sistemáticamente. Por otra parte, T1 muestra que la fertilización anual sólo tiene efectos positivos iniciales en L (figura 4a). La biomasa fustal incrementa sistemáticamente en ambos tratamientos (figura 4b), presentando T2 el mayor incremento después del último raleo con 79,8 t ha1 en un período de 5 años. En cambio, T1 muestra un incremento muy similar en biomasa después del raleo con 76,5 t ha1, para el mismo período. Estos resultados sugieren que el efecto de la fertilización en la biomasa fustal es limitado por la estructura de copas. Los valores de incremento medio anual (IMA) estimados y simulados (cuadro 4) son similares a los valores reportados por Toro y Gessel (1998), para zonas costeras de suelos granítico-metamórficos, de la VI Región de Chile. Sin embargo, los valores obtenidos en los distintos rodales, 17,5 m3 ha1 año1 en T1 y 19,3 m3 ha1 año1 en T2, son ilustrativos respecto del efecto del régimen silvícola y el manejo de los recursos del sitio en la productividad. En efecto, el mayor IMA de T2 con respecto a T1 se atribuye a la mayor densidad residual. Asimismo, estos resultados muestran la influencia de la densidad en el Dap, puesto que T2 presenta valores aproximadamente un 38,5% inferiores que los Dap medios de T1 (cuadro 4).
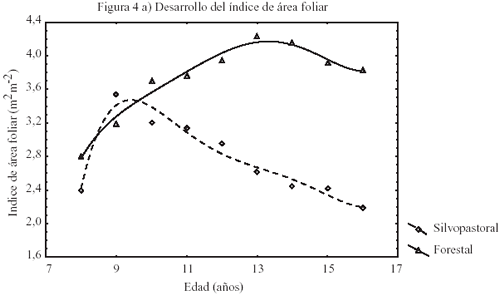
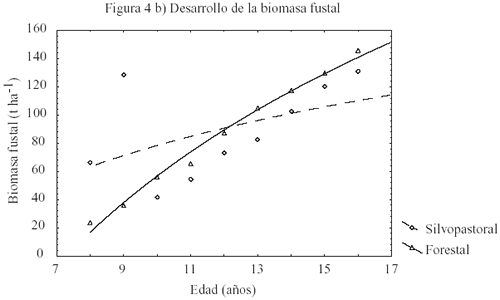
Figura
4: Diagrama de dispersión para el efecto del raleo en el
desarrollo del índice de área foliar (m2 m2),
biomasa fustal (t ha1), relación entre el índice de
área foliar con la biomasa fustal y relación entre la transpiración
(mm) y el índice de área foliar en pino radiata de 16 años. Dispersion diagram of the effect of thinning on the development of leaf area index (m2/m2), stem biomass (t/ha), the relationship between leaf area index and stem biomass, and the relationship between transpiration (mm) and leaf area index, in 16-year-old radiata pine. |
Valores simulados
y observados para el Dap, volumen, incremento medio anual y simulados
para el máximo índice de área foliar y la proporción de agua utilizada
en transpiración con la precipitación en pino radiata de 16 años.
Simulated and observed values for diameter at breast height (DBH), volume, and annual mean increment, and simulated values for maximum leaf area index and the water proportion used in transpiration with precipitation, for 16-year-old radiata pine. |
| - | Regímenes
de manejo |
|
T1 |
T2 |
|
| Proporción Transpiración/Precipitación Ranking de fertilidad del suelo Máximo Indice de área foliar (m2 m2) IMA Simulado (m3 ha1 año1) IMA Observado (m3 ha1 año1) Dap Simulado (cm) Dap Observado (cm) |
0,57 1,0 3,5 17,4 17,5 41,1 43,4 |
0,61 1,0 4,2 19,3 19,3 28,6 29,8 |
| Nota: Máxima capacidad de agua del suelo, 125 (mm); el quantum de eficiencia del dosel fue modificado por el déficit de presión de vapor, los días de heladas y el ranking de fertilidad (en este ranking los rodales fertilizados anualmente se le asignó el valor 1 y el no fertilizado un valor de 0,1). |
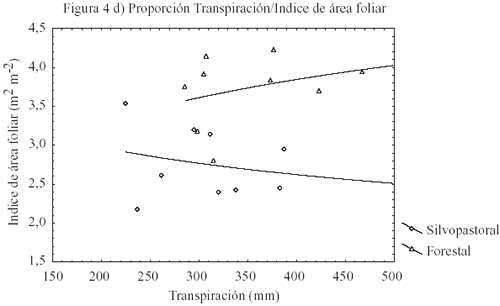
Los valores de transpiración (cuadro 4) están en el orden de lo reportado por Teskey y Sheriff (1996). En general, se observa que la densidad final y el índice de área foliar son los factores determinantes en el consumo hídrico a nivel de rodal (figura 4d). En zonas semiáridas y suelos graníticos y metamórficos erosionados, como es el caso de la zona de estudio, el agua es la principal limitante de la productividad. Sin embargo, la modelación indica que 125 mm de agua disponible en el suelo son suficientes para incrementar la productividad del bosque.
DISCUSION
Validación del modelo en la predicción del crecimiento en volumen. El modelo 3-PG es un buen predictor del volumen con estimaciones confiables, bajo error y libres de sesgo (cuadro 3). Por ello, su empleo para el análisis de la especie en estudio resultó altamente valioso, puesto que proporcionó antecedentes que no aportan los modelos estadísticos tradicionales (cuadro 4).
Efecto del régimen silvícola
y el raleo en el desarrollo del índice de área foliar y la productividad.
El desarrollo de L en el régimen de manejo forestal, en comparación al
rodal con manejo silvopastoral, resultó marcadamente diferente (figura
3 a). Al respecto, se espera que una distribución homogénea de los
árboles en el espacio, como es el caso de T2, logre alcanzar el potencial
de L y permita maximizar la intercepción de ![]() p.a.u,
(Grace et al. 1987). Sin embargo, en rodales con manejo
silvopastoral, el diseño de plantación y el régimen de fertilización permitieron
que los rodales alcanzaran rápidamente la ocupación parcial del sitio.
Estos resultados concuerdan con lo postulado por Vose y Allen (1988) en
el sentido de que el desarrollo y el potencial de L es altamente dependiente
de la densidad del rodal, tamaño del árbol y la oferta de los recursos
del sitio. En este estudio, la estructura de copas y la densidad después
del raleo impidieron alcanzar el máximo potencial de L, aunque la fertilidad
del sitio fue mejorada.
p.a.u,
(Grace et al. 1987). Sin embargo, en rodales con manejo
silvopastoral, el diseño de plantación y el régimen de fertilización permitieron
que los rodales alcanzaran rápidamente la ocupación parcial del sitio.
Estos resultados concuerdan con lo postulado por Vose y Allen (1988) en
el sentido de que el desarrollo y el potencial de L es altamente dependiente
de la densidad del rodal, tamaño del árbol y la oferta de los recursos
del sitio. En este estudio, la estructura de copas y la densidad después
del raleo impidieron alcanzar el máximo potencial de L, aunque la fertilidad
del sitio fue mejorada.
Ambos tratamientos incrementaron
la biomasa fustal después del raleo, aunque el rodal con manejo forestal
mostró un comportamiento diferente en el desarrollo de la biomasa fustal
que el rodal silvopastoral en relación al incremento de L (figura
4c). Al respecto, se ha considerado que el incremento de la biomasa
fustal es función de la eficiencia de crecimiento y L (Mäkelä 2000b).
Además, existe una directa relación de ![]() p.a.u,
con la producción de materia seca (Grace et al. 1987).
Por tanto, se espera que el raleo tenga efectos positivos en el incremento
de L y en la eficiencia de crecimiento, lo que explica la mayor biomasa
fustal (Waring y Runing 1998). El raleo en T2 efectivamente incrementó
L y la eficiencia de crecimiento. Sin embargo, como se desprende de la
figura 3c, en el rodal con manejo silvopastoral el raleo tuvo sólo un
efecto inicial. En rodales manejados, la hipótesis de McMurtrie et
al. (1992) es que la fertilización incrementa L y la biomasa
fustal y que L sólo es limitado por condiciones de agua y nutrientes.
Sin embargo, esta hipótesis no se verificó en el rodal con manejo silvopastoral,
ya que el límite de L (cuadro 4) estuvo dado por la
estructura de copas debido al diseño de plantación y la baja densidad
residual de los rodales.
p.a.u,
con la producción de materia seca (Grace et al. 1987).
Por tanto, se espera que el raleo tenga efectos positivos en el incremento
de L y en la eficiencia de crecimiento, lo que explica la mayor biomasa
fustal (Waring y Runing 1998). El raleo en T2 efectivamente incrementó
L y la eficiencia de crecimiento. Sin embargo, como se desprende de la
figura 3c, en el rodal con manejo silvopastoral el raleo tuvo sólo un
efecto inicial. En rodales manejados, la hipótesis de McMurtrie et
al. (1992) es que la fertilización incrementa L y la biomasa
fustal y que L sólo es limitado por condiciones de agua y nutrientes.
Sin embargo, esta hipótesis no se verificó en el rodal con manejo silvopastoral,
ya que el límite de L (cuadro 4) estuvo dado por la
estructura de copas debido al diseño de plantación y la baja densidad
residual de los rodales.
Por tanto, los resultados obtenidos sugieren que en pino radiata establecido en suelos pobres los factores determinantes en el incremento de la biomasa fustal y el IMA son la masa foliar del dosel, la estructura de las copas (Perry 1985, Cannel 1989) y la densidad final (Vose y Allen 1988, Landsberg y Gower 1997), más que los recursos del sitio. Los antecedentes del régimen silvopastoral sugieren que la fertilización tiene efectos en el desarrollo de L, cuyo límite está dado además por la estructura de copas y la densidad del rodal. Por ello, el efecto de la fertilización en el incremento de biomasa fustal podría explicarse, además, por un cambio en la localización de la biomasa de raíces finas a crecimiento fustal debido al régimen de fertilización (Santantonio y Santantonio 1987, Beets y Whitehead 1996, Rodríguez et al. 2001) y al menor consumo hídrico del régimen silvopastoral (Waring y Schlesinger 1985, Waring 1987, Oliver y Larson 1990, Waring y Runing 1997).
La idea generalizada es que en zonas semiáridas un incremento en productividad implica un mayor consumo hídrico. En este estudio, la máxima cantidad de agua disponible en el suelo (cuadro 4) fue la misma en ambos tratamientos. Sin embargo, los valores de IMA a los 16 años fueron mayores en el régimen de manejo forestal que en el silvopastoral. De acuerdo con los resultados de la simulación, el rodal silvopastoral consumió menos agua que el rodal con manejo forestal. Al respecto, Eastham et al. (1990) indican que la productividad y uso del agua entre árboles y praderas está regulada por la densidad de los árboles y no por el tamaño de los árboles (cuadro 3, figura 4d). Ello concuerda con la hipótesis de Teskey y Sheriff (1996) en que árboles grandes extraen agua del mismo volumen de suelo que árboles pequeños y que el consumo hídrico en rodales de pino radiata está altamente correlacionado con la disponibilidad de agua en el primer metro del perfil del suelo. Además, la eficiencia en el uso del agua se atribuye a la disminución en la tasa de recambio de raíces finas, como fue el caso del rodal con manejo silvopastoral.
CONCLUSIONES
El desarrollo y valores máximos del índice de área foliar se presentaron fuertemente controlados por el diseño de plantación y la densidad del rodal. La fertilización tiene efectos positivos sobre el índice de área foliar, en la medida que no sea limitado por la estructura de copas. Por la fuerte relación entre la radiación fotosintéticamente activa con la producción de materia seca (Grace et al. 1987), la ocupación del sitio por las copas explica la magnitud en la producción de madera (Vose y Allen 1987) en el rodal con manejo forestal. Cuando se alcanza el máximo L, los rodales incrementan la biomasa fustal en forma transitoria, a expensas de la biomasa de raíces finas por efecto del régimen de fertilización (Santantonio y Santantonio 1987, Beets y Whitehead 1996), aunque limitada por el agua. El raleo implica una perturbación en el desarrollo del rodal, por su efecto en la estructura de copas y la disponibilidad de los recursos del sitio afectando la producción final de madera. Por consiguiente, la maximización de la producción de madera dependerá del manejo conjunto de los recursos del sitio con la intensidad y oportunidad del raleo (Snowdon y Benson 1992). Para ello, la simulación del crecimiento mediante modelos basados en procesos es una herramienta necesaria. Sin embargo, es importante incorporar a la modelación el raleo, puesto que la ley de autorraleo opera en plazos mayores que las rotaciones usuales de plantaciones artificiales en especies de rápido crecimiento.
AGRADECIMIENTOS
A la Corporación Nacional Forestal por su valiosa contribución financiera y su apoyo logístico. Al Ministerio de Planificación y Cooperación (MIDEPLAN). Al Ingeniero Forestal de CONAF Sr. César Cabrera y a los trabajadores del Centro Experimental Forestal TANUME, por su valioso apoyo logístico.
BIBLIOGRAFIA
BEETS, P. N. and D. WHITEHEAD. 1996. Carbon partitioning in Pinus radiata stands in relation to leaf nitrogen status, Tree Physiology. 16: 131-138.
BRISTOW, K. L. and G. S. CAMPBELL. 1984. On the relationship between incoming solar radiation and daily maximun and minimun temperature, Agricultural Forest Meteorology, 32: 159-166.
CANNEL, M. G. R. 1989. Physiological basis of wood production: a review, Scandinavian Journal of Forest Research. 4: 459-490.
CARLYLE, C. 1995. Managing site resources to maximize the productivity of Pinus radiata plantations. In: X Silvotecna-Expocorma. Concepción, Chile.
COOPS, N. C., R. H. WARING, and J. J. LANDSBERG. 1998. Assesing forest productivity in Australia and New Zealand using a physiologically-based model driven with averaged monthly weather data and satellite-derived estimates of canopy photosynthetic capacity, Forest Ecology and Management 104:113-127.
EASTHAM, J., C. W. ROSE, D. A. CHARLES-EDWARDS, D. M. CAMERON and S. J. RANCE. 1990. Planting density effects on water use efficiency of trees and pasture in an agroforestry experiment. New Zealand Journal of Forest Science. 1: 39-53.
GRACE, J., P. G. JARVIS and J. M. NORMAN. 1987. Modelling the interception of solar radiant energy in intensively managed stand, New Zealand Journal of Forest Science. 17(2/3): 193-209.
HUSCH, B., C. MILLER and T. BEERS. 1982. Forest mensuration. Krieger Publishing. New York, USA. 402 p.
LANDSBERG, J. J. and F. J. HINGSTON. 1996. Evaluating a simple radiation/dry matter conversion model using data from Eucalyptus globulus plantations in western Australia. Tree Physiology. 16: 801-808.
LANDSBERG, J. J. and S. T. GOWER. 1997. Applications of physiological ecology to forest management. Academic Press, San Diego, USA, 354 p.
LANDSBERG, J. J. and R. H. WARING. 1997. A generalised model of forest productivity using concepts of radiationuse efficiency, carbon balance and partitioning, Forest Ecology and Management. 95: 209-228.
McMURTRIE, R. E., D. A. ROOK and F. M. KELLIHER. 1990. Modelling the yield of Pinus radiata on a site limited by water and nitrogen, Forest Ecology and Management 30: 381-413.
MÄKELÄ, A., J. J. LANDSBERG, A. R. EK, T. E. BURK, M. TER-MIKAELIAN, G. I. AGREN, C. D. OLIVER and P. PUTTONEN. 2000a. Process-based model for forest ecosystem management: current state of the art and challenges for practical implementations. Tree Physiology. 20: 289-298.
MÄKELÄ, A., R. SIEVÄNEN, M. LINDNER and P. LASCH. 2000b. Application of volume growth and survival graphs in the evaluation of four process-based models, Tree Physiology. 20: 347-355.
McMURTRIE, R. E. and J. J. LANDSBERG. 1992. Using a simulation model to evaluate the effects of water and nutrients on the growth and carbon partitioning of Pinus radiata, Forest Ecology and Management. 52: 243-260.
McMURTRIE, R. E., D. A. ROOK and F. M. KELLIHER. 1992. Modelling the yield of Pinus radiata on a site limited by water and nitrogen, Forest Ecology and Management. 30: 381-413.
MENCUCCINI, M. And J. GRACE. 1996. Hydraulic conductance, light interception and needle nutrient concentration in Scots pine stand and their relation with net primary productivity, Tree Physiology. 16: 459-468.
OLIVER, C. D. and B. C. LARSON. 1996. Forest stand dynamics. John Wiley & Sons, New York, USA, 520 p.
PERRY, D. A. 1985. The competition process in forest stand. pp. 481-506. In M.G.R. CANNEL and J. E. JACKSON (editors). Attributes of trees as Crop Plants. Institute of Terrestrial Ecology, Penicuick, UK.
RAISON, R. J., P. K. KHANNA, M. J. CONNEL and R. A. FALKINER. 1992. Effects of water availability and fertilization on nitrogen cycling in a stand of Pinus radiata. Forest Ecology and Management. 30: 31-43.
ROBERTS, S. D., J. N. LONG and F. W. SMITH. 1993. Canopy stratification and leaf area efficiency: a conceptualization, Forest Ecology and Management. 60: 143-156.
RODRIGUEZ, R., G. HOFFMANN, M. A. ESPINOSA AND D. RIOS. 2001. Comparative study of biomass and leaf area among silvopastoral regimes and a traditional forestry regime in radiata pine. Submitted to Revista Chilena de Historia Natural.
RUNING, S. W., R. R. NEMANI and R. D. HUNGENFORD. 1987. Extrapolation of synoptic meteorological data in mountains terrain and its use for simulating forest evapotranspiration and photosynthesis. Canadian Journal of Forest Research. 17: 472-483.
RUNING, S. W. and S. T. GOWER. 1991. Forest-BGC, a general model of forest ecosystem process for regional applications II. Dynamic carbon allocations and nitrogen budgets. Tree Physiology. 9: 147-160.
SANDS, P. 2000. 3PGpjs a user-friendly interface to 3-PG, the Landsberg and Waring model of forest productivity. Cooperative Research Centre for Sustainable Production Forestry and CSIRO Forestry and Forest Products. Technical Report 29. Tasmania, Australia. 16 p.
SANTANTONIO, D. 1989. Dry-matter partitioning and fineroot production in forests-new approaches to a difficult problem. pp. 57-72. In J.S. PEREIRA and R. H. WARING (editors). Biomass Production by Fast-Growing Trees. Kluwer Academic. Dordrecht, Netherlands.
SANTANTONIO, D. and E. SANTANTONIO. 1987. Seasonal changes and dead fine roots during two succesive years in a thinned plantation of Pinus radiata in New Zealand, New Zealand Journal of Forest Science. 17 (2/3): 315-328.
SNOWDON, P. and M. L. BENSON. 1992. Effects of combinations of irrigation and fertilisation on the growth and above-ground biomass production of Pinus radiata, Forest Ecology and Management. 52: 87-116.
TESKEY, R. O. and D. W. SHERIFF. 1996. Water use by Pinus radiata trees plantation, Tree Physiology. 19: 273- 279.
TORO, J. and S. GESSEL. 1998. Radiata pine plantations in Chile, New Forest. 00: 1-12.
VAN LAAR, A. and A. AKÇA. 1997. Forest mensuration. Cuvillier Verlag, Göttingen, Germany, 418 p.
VOSE, J. M. and H. L. ALLEN. 1988. Leaf area, stemwood growth, and nutrition relationships in loblolly pine, Forest Science. 34: 547-563.
WANG, Y. P. and P. G. JARVIS. 1990. Influence of crown properties on PAR absortion, photosynthesis, and transpiration in sitka spruce: Application of a model (MAESTRO), Tree Physiology. 7: 297-316.
WARING, R. H. 1987. Characteristics of trees predisposed to die, BioScience 37: 569-574.
WARING, R. H. 2000. A process model analysis of environmental limitations on the growth of sitka spruce plantations in Great Britain, Forestry 73 (1): 65-79.
WARING, R. H. and W. H. SCHLESINGER. 1985. Forest Ecosystem. Academic Press, New York, USA, 339 p.
WARING, R. H., J. J. LANDSBERG and M. WILLIAMS. 1998. Net primary production of forest: a constant fraction of gross primary production?, Tree Physiology. 18: 129- 134.
WARING, R. H. and S. W. RUNING. 1998. Forest ecosystem. Analysis at multiples scales. Academic Press, San Diego, USA, 370 p.
Recibido: 16.11.2001
Aceptado: 04.09.2002